1. Общие свойства возбудимых тканей.
Возбудимость – способность ткани отвечать на раздражение изменением ряда своих свойств. Показатель возбудимости – порог раздражения . Это минимальное по силе раздражение, способное вызвать видимую ответную реакцию ткани.
Проводимость – способность ткани проводить возбуждение по всей своей длине. Показатель проводимости – скорость проведения возбуждения. Проводимость напрямую зависит от возбудимости ткани: чем выше возбудимость, тем выше проводимость, так как быстрее возбуждается расположенный рядом участок ткани.
Рефрактерность – способность ткани терять или снижать возбудимость в процессе возбуждения. При этом в ходе ответной реакции ткань перестает воспринимать раздражитель. Рефрактерность бывает абсолютной (нет ответа ни на какой раздражитель) и относительной (возбудимость восстанавливается, и ткань отвечает на подпороговый или сверхпороговый раздражитель). Показатель рефрактерности (рефрактерный период ) — время, в течение которого возбудимость ткани снижена. Рефрактерный период тем короче, чем выше возбудимость ткани
Лабильность – способность возбудимой ткани реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом волн возбуждения, возникающих в ткани в единицу времени (1 с) в точном соответствии с ритмом наносимых раздражений без явления трансформации. Лабильность определяется продолжительностью рефрактерного периода (чем короче рефрактерный период, тем больше лабильность).
Для мышечной ткани характерна такжесократимость . Сократимость – способность мышцы отвечать сокращением на раздражение.
2.Классификация раздражителей
Раздражитель – фактор, способный вызвать ответную реакцию возбудимых тканей.
1) естественные (нервные импульсы, возникающие в нервных клетках и различных рецепторах);
2) искусственные: физические (механические – удар, укол; температурные – тепло, холод; электрический ток – переменный или постоянный), химические (кислоты, основания, эфиры и т. п.), физико-химические (осмотические – кристаллик хлорида натрия).
По своей природе раздражители бывают:
- химические;
- физические;
- механические;
- термические;
- биологические.
По биологическому соответствию , то есть насколько раздражитель соответствует данной ткани.
1) адекватные – раздражители, которые соответствуют данной ткани . Например, для сетчатки глаза свет – все остальные раздражители не соответствуют сетчатке, для мышечной ткани – нервный импульс и т.д.;
2) неадекватные – раздражители, которые не соответствуют данной ткани . Для сетчатки глаза все раздражители кроме светового будут неадекватные, а для мышечной ткани все раздражители, кроме нервного импульса.
По силе :
1) подпороговые раздражители – это сила раздражителя при которой не возникает ответная реакция;
2) пороговыйраздражитель – это минимальная сила, которая вызывает ответную реакцию при бесконечном времени действия. Эту силу еще называют реобазой – она единственная для каждой ткани;
3) надпороговые , или субмаксимальные ;
4)максимальный раздражитель – это минимальная сила при которой возникает максимальная ответная реакция ткани ;
5) сверхмаксимальные раздражители – при этих раздражителях реакция ткани либо максимальная, либо уменьшается, либо временно исчезает.
Таким образом, для каждой ткани существует один пороговый раздражитель , один максимальный и множество подпороговых, надпороговых и сверхмаксимальных.
3. Физиология клеточных мембран. Механизмы трансмембранного транспорта.
— Пограничная функция. Мембрана отграничивает цитоплазму от межклеточной жидкости, а большинство внутриклеточных структур: митохондрии, ядро, эндоплазматическую сеть – от цитоплазмы.
— Биотрансформирующая функция. Любое вещество, проходя через мембрану, вступает с ней в сложное взаимодействие и претерпевает ряд биохимических превращений. В результате биотрансформации лекарственное вещество, как правило, переходит в форму, легко усвояемую клеткой.
— Транспортная функция. Перенос веществ через биологические мембраны связан с процессами метаболизма, поддержанием постоянства внутренней среды клетки, возбуждением и проведением нервного импульса.
Существует два основных типа переноса: пассивный (фильтрация, диффузия, облегченная диффузия, осмос) и активный (работа мембранных белковых «насосов»)
Пассивный транспорт. Фильтрация осуществляется через мембранные белковые каналы – поры, зависит от разности давлений снаружи и внутри клетки и проницаемости мембраны для жидкости и низкомолекулярных веществ. Диаметр пор чрезвычайно мал, поэтому фильтруются только низкомолекулярные вещества, вода и некоторые ионы.
Диффузия — пассивное передвижение молекул или ионов по градиенту концентрации (из области высокой концентрации в область низкой). Осмос представляет собой частный случай диффузии растворителя через полупроницаемую мембрану, не пропускающую растворенные вещества.
Пассивный транспорт не требует затрат энергии.
Активный транспорт. Это универсальный для всех видов мембран перенос веществ против концентрационных или электрохимических градиентов (из области низкой концентрации в область высокой). При помощи активного транспорта переносятся гидрофильные полимерные молекулы, неорганические ионы (Na, Ca, K) , водород, сахара, аминокислоты, витамины, гормоны и лекарственные вещества. Активный транспорт осуществляется с обязательной затратой энергии, образующейся при расщеплении (окислительное фосфорилирование) аденозинтрифосфорной кислоты (АТФ).
Разновидностью активного транспорта, связанной с деятельностью самой клетки, является микровезикулярный транспорт (пиноцитоз, экзоцитоз и фагоцитоз). При пиноцитозе происходит активное поглощение клеткой жидкости из окружающей среды с формированием пузырьков и последующим переносом их через цитоплазму. Процесс слияния пузырьков с мембраной клетки и выделение клеткой вещества в виде секреторных гранул или вакуолей называется экзоцитозом . Явление фагоцитоза заключается в способности клеток активно захватывать и поглощать микроорганизмы, разрушенные клетки и инородные частицы.
— Рецепторная функция. Биологические мембраны имеют большое количество рецепторов – участков, молекулярная структура которых характеризуется избирательным сродством к определенным физиологически активным веществам: гормонам, медиаторам, антигенам.
— Образование межклеточных контактов.
— Генерация биоэлектрических потенциалов. В ходе эволюции у железистого эпителия, мышечной и нервной тканей появилось свойство возбудимости – способность реагировать на воздействие окружающей среды возбуждением. Внешним проявлением возбуждения является возникновение биоэлектрического потенциала.
4. Ионные механизмы мембранного потенциала покоя
О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма, нет видимого функционального отправления ткани.
Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин:
1) неодинакового распределения ионов по обе стороны мембраны. Внутри клетки находится больше всего ионов К, снаружи его мало. Ионов Na и ионов Cl больше снаружи, чем внутри. Такое распределение ионов называется ионной асимметрией;
2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ.
За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концентрации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил:
1) силы диффузии;
2) силы электростатического взаимодействия.
Значение электрохимического равновесия:
1) поддержание ионной асимметрии;
2) поддержание величины мембранного потенциала на постоянном уровне.
В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентрационно-электрохимическим.
Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ. Работа натрий-калиевого насоса обеспечивает:
1) высокую концентрацию ионов К внутри клетки, т. е. постоянную величину потенциала покоя;
2) низкую концентрацию ионов Na внутри клетки, т. е. сохраняет нормальную осмолярность и объем клетки, создает базу для генерации потенциала действия;
3) стабильный концетрационный градиент ионов Na, способствуя транспорту аминокислот и сахаров.
5. Мембранный потенциал действия: фазы, ионные механизмы.
Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный.
Возбуждение нервной клетки под действием химического сигнала (реже электрического импульса) приводит к возникновению потенциала действия . Это означает, что потенциал покоя -60 мВ скачком изменяется на +30 мВ и спустя 1 мс принимает исходное значение. Процесс начинается с открывания Nа+-канала (1). Ионы Na+устремляются в клетку (по градиенту концентрации), что вызывает локальное обращение знака мембранного потенциала (2). При этом Na+-каналы тотчас закрываются, т. е. поток ионов Na+ в клетку длится очень короткое время (3). В связи с изменением мембранного потенциала открываются (на несколько мс) потенциал-управляемые К+-каналы (2) и ионы К+ устремляются в обратном направлении, из клетки. В результате мембранный потенциалпринимает первоначальное значение (3), и даже превышает на короткое времяпотенциал покоя (4). После этогонервная клетка вновь становится возбудимой.
За один импульс через мембрану проходит небольшая часть ионов Na+ и К+, и концентрационные градиенты обоихионов сохраняются (в клетке выше уровень К+, а вне клетки выше уровень Na+). Поэтому по мере получения клеткойновых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращениемембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.
Восходящая ветвь графика:
- потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ);
- нарастающий локальный потенциал – пропорциональная раздражителю деполяризация;
- критический уровень деполяризации (–50 мВ) – резкое ускорение деполяризации (за счет самораскрытия натриевых каналов), с этой точки начинается спайк – высокоамплитудная часть потенциала действия;
- самоусиливающаяся круто нарастающая деполяризация;
- переход нулевой отметки (0 мВ) – смена полярности мембраны;
- «овершут» – положительная поляризация (инверсия , или реверсия, заряда мембраны);
- пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия.
Нисходящая ветвь графика:
- реполяризация – восстановление прежней электроотрицательности мембраны;
- переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную;
- переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости;
- следовые процессы (следовая деполяризация или следовая гиперполяризация);
- восстановление потенциала покоя – норма (–70 мВ).

6. Классификация нервных волокон.
А – нервные волокна с самой толстой миелиновой оболочкой. Наиболее высокая скорость передачи нервного импульса.
В – миелиновая оболочка тоньше, скорость проведения возбуждения ниже
С – безмиелиновые волокна с относительно низкой скоростью передачи импульса.
| Тип волокна |
Диаметр волокна (мк) |
Скорость проведения (м/сек) |
Длительность потенциала действия (мсек) |
Длительность отрицательного следового потенциала (мсек) |
Длительность положительного следового потенциала (мсек) |
Функция |
| А (α) | 12-22 | 70-120 | 0,4-0,5 | 12-20 | 40-60 | Моторные волокна скелетных мышц, афферентные волокна мышечных рецепторов |
| А (β) | 8-12 | 40-70 | 0,4-0,6 | — | — | Афферентные волокна от рецепторов прикосновения |
| А (γ) | 4-8 | 15-40 | 0,5-0,7 | — | — | Афферентные волокна от рецепторов прикосновения и давления, эфферентные волокна к мышечным веретенам |
| А (Δ) | 1-4 | 5-15 | 0,6-1,0 | — | — | Афферентные волокна от некоторых рецепторов тепла, давления, боли |
| В | 1-3 | 3-14 | 1-2 | Отсут- ствует |
100-300 | Преганглионарные вегетативные волокна |
| С | 0,5-1,0 | 0,5-2 | 2,0 | 50-80 | 300-1000 | Преганглионарные вегетативные волокна, афеерентные волокна от некоторых рецепторов боли, давления, тепла |
Возбуждение. Возбудимость. Проводимость. Рефрактерность и лабильность. Физиологические свойства нервных волокон (безмиелиновых и миелиновых). Утомляемость нервного волокна. Физиологические свойства синапсов.
"Все регулируется, течет по расчищенным руслам, совершает свой кругооборот в соответствии с законом и под его защитой".
И. Ильф и Е. Петров "Золотой теленок"
Все клетки и ткани живого организма под действием раздражителей переходят из состояния относительного физиологического покоя в состояние активности (возбуждения). Наибольшая степень активности наблюдается в нервной и мышечной ткани.
Главными свойствами возбудимых тканей являются: I. возбудимость, II проводимость, III рефрактерность и лабильность, которые связаны с одним из самых общих свойств живого — раздражимостью.
Изменения в окружающей среде или организме называют раздражителями, а их действие — раздражением.
По природе раздражители бывают: механические, химические, электрические, температурные.
По биологическому признаку раздражители делятся на:
адекватные, которые воспринимаются соответствующими специализированными рецепторами (глаза — свет, уха — звук, кожа — боль, температура, прикосновение, давление, вибрация);
2. неадекватные, к которым специализированные рецепторы не приспособлены, но воспринимают их при чрезмерной силе и длительности (удар — глаз — свет).
Наиболее общим, адекватным и естественным раздражителем для всех клеток и тканей организма является нервный импульс.
Основные физиологические свойства нервной ткани (возбудимость, проводимость, рефрактерность и лабильность) характеризуют функциональное состояние нервной системы человека, определяют его психические процессы.
I. Возбудимость — способность живой ткани отвечать на действие раздражителя возникновением процесса возбуждения с изменением физиологических свойств.
Колличественной мерой возбудимости является порог возбуждения, т.е. минимальная величина раздражителя, способная вызвать ответную реакцию тканей.
Раздражитель меньшей силы называют подпороговым, а большей — надпороговым.
Возбудимость представляет собой, в первую очередь, изменение обмена веществ в клетках тканей. Изменение обмена веществ сопровождается переходом через клеточную мембрану отрицательно и положительно заряженных ионов, которые изменяют электрическую активность клетки. Разность потенциалов в покое между внутренним содержимым клетки и клеточной оболочкой, составляющая 50-70 мВ (миллиВольт) называется мембранным потенциалом покоя.
Основой этого состояния клетки является избирательная проницаемость мембраны по отношению к ионам К+ и Na+. Ионам Na+, находящимся во внеклеточной среде, через мембрану в клетки путь закрыт, а К+ свободно проникает через поры клеточной мембраны из цитоплазмы клетки в тканевую жидкость. В результате в цитоплазме остаются отрицательно заряженные ионы, а на поверхности мембраны накапливаются положительно заряженные ионы К+ и Na+.
При возбуждении клетки проницаемость ионов Na+ резко увеличивается, и они устремляются в цитоплазму, снижая потенциал покоя до нуля, а затем увеличивая разность потенциалов противоположного значения до 80-110 мВ. Такое кратковременное (0,004-0,005 сек) изменение разности потенциалов называется потенциалом действия (спайком); англ. spike — острие.
Вслед за этим нарушенное равновесие ионов вновь восстанавливается. Для этого существует специальный клеточный механизм — "натрий-калиевый насос", который обеспечивает активное "выкачивание" Na+ из клетки и «нагнетание» в нее К+. Таким образом, существуют 2 типа движения ионов через клеточную мембрану:
1 — пассивный ионный транспорт по градиенту концентрации ионов;
2 — активный ионный транспорт против градиента концентрации, осуществляемый "натрий-калиевым насосом" с затратой энергии АТФ.
Вывод: возбуждение нервной клетки связано с изменением обмена веществ и сопровождается появлением электрических потенциалов (нервных импульсов).
Проводимость — способность живой ткани проводить волны возбуждения — биоэлектрические импульсы.
Для обеспечения гомеостатического единства все структуры организма (клетки, ткани, органы и т.д.) должны иметь возможность пространственного взаимодействия. Распространение возбуждения от места его возникновения до исполнительных органов — один из основных способов такого взаимодействия. Возникший в месте нанесения раздражения потенциал действия является причиной раздражения соседних, невозбужденных участков нервного (или мышечного) волокна. Благодаря этому явлению волна потенциала действия создает ток действия, который распространяется по всей длине нервного волокна. В безмиелиновых нервных волокнах возбуждение проводится с некоторым затуханием — декрементом, а в миелиновых нервных волокнах — без затухания. Проведение возбуждения также сопровождается изменением обмена веществ и энергии.
III. Рефрактерность — временное снижение возбудимости ткани, возникающее при появлении потенциала действия. В этот момент повторные раздражения не вызывают ответной реакции (абсолютная рефрактерность). Она длится не более 0,4 миллисекунды, а затем наступает фаза относительной рефракторности, когда раздражение может вызвать слабую реакцию. Эта фаза сменяется фазой повышенной возбудимости — супернормальности.
Такая динамика возбудимости обусловлена процессами изменения и восстановления равновесия ионов на мембране клетки.
Профессор Н.Е. Введенский исследовал особенности этих процессов и установил, что возбудимые ткани могут отвечать разным числом потенциалов действия на определенную частоту раздражений. Он назвал это явление лабильностью (функциональной подвижностью).
Лабильность — свойство возбудимой ткани воспроизводить максимальное число потенциалов действия в единицу времени.
Максимальная лабильность — у нервной ткани. Частота раздражений, вызывающая максимальную реакцию называется оптимальной (лат. optimum — наилучший), а вызывающая угнетение реакции — пессимальной (лат. pessimum — наихудший).
*Нервное волокно — до 1000 имп./сек, мышца — 200-250 имп./сек., синапс — до 100-125 имп./сек.
Пессимум — активная реакция ткани, направленная на защиту ее от чрезмерных раздражений. Это одна из форм проявления торможения. Возбуждение и торможение это противоположные по значению саморегулирующиеся процессы, которые устанавливают "золотую середину" уровня отношений организма со средой.
Нервные волокна (отростки нервных клеток) обладают всеми свойствами возбудимых тканей, а проведение нервных импульсов является их специальной функцией. Скорость проведения возбуждения зависит от:
1 — диаметра волокон (толще ® быстрее),
2 — строения их оболочки.
Безмиелиновые (безмякотные) волокна покрыты только леммоцитами (шванновскими клетками). Между ними и осевым цилиндром (аксоном нейрона) имеется щель с межклеточной жидкостью, поэтому, клеточная мембрана остается неизолированной. Импульс распространяется по волокну со скоростью всего 1-3 м/сек.
Миелиновые волокна покрыты спиральными слоями шванновских клеток с прослойкой миелина — жироподобного вещества с высоким удельным сопротивлением. Миелиновая оболочка через промежутки равной длины прерывается, оставляя оголенными участки осевого цилиндра длиной » 1 мкм.
Из-за такого строения электрические токи могут входить в волокна и выходить из них только в области неизолированных участков — перехватов Ранвье. При нанесении раздражения в ближайшем перехвате возникает деполяризация, а соседние перехваты поляризованы. Между ними возникает разность потенциалов, которая приводит к появлению круговых токов действия.
Таким образом, импульс в миелиновом волокне проходит скачкообразно (сальтаторно) от перехвата к перехвату. Возбуждение при этом распространяется без затухания, а скорость проведения импульса достигает 120-130 м/сек.
При нанесении раздражения на нервное волокно происходит двустороннее распространение возбуждения — в центростремительном и центробежном направлении. Это не противоречит принципу одностороннего проведения импульсов, и объясняется первичностью появления возбуждения в рецепторах или нервных центрах, а также наличием синапсов. Нейротрансмиттер (медиатор) содержится только в пресинаптическом аппарате и переносит потенциал только однонаправленно (см. лекцию по анатомии № 2).
Возбуждение проводится не только в нужном направлении, но и по одному изолированному волокну, не распространяясь на соседние волокна. Это обуславливает строго координированную рефлекторную деятельность. Например, седалищный нерв диаметром до 12 мм несет в себе тысячи нервных волокон (миелиновых и безмиелиновых, чувствительных и двигательных, соматических и вегетативных). В случае неизолированного проведения возбуждения наблюдалась бы хаотическая ответная реакция.
Изолированное проведение возбуждения в миелиновых волокнах обеспечивается миелиновой оболочкой, а в безмиелиновых — высоким удельным сопротивлением окружающей межклеточной жидкости (отсюда и затухание потенциала).
Н.Е. Введенский в 1883 году впервые установил, что нерв малоутомляем. Малая утомляемость нервных волокон объясняется тем, что энергетические затраты в них при возбуждении незначительны, а процессы восстановления протекают быстро. В организме нервные волокна работают также с недогрузкой. Например, двигательное волокно высоколабильно и может проводить до 2500 имп./сек. Из нервных же центров поступает не более 50-40 имп./сек.
Вывод: практическая неутомляемость нервных волокон связана с небольшими энергетическими затратами, с высокой лабильностью нервных волокон, с постоянной недогрузкой волокон.
Синапсы (см. строение в лекции по анатомии № 2) обладают следующими физиологическими свойствами:
1 — одностороннее проведение возбуждения, которое связано с особенностями строения самого синапса,
Поиск Лекций
Физиологические свойства скелетных мышц. Фазовые изменения возбудимости нервной и мышечной ткани. Методы измерения возбудимости
Физиологические свойства мышц
Возбудимость — способность приходить в состояние возбуждения при действии раздражителей.
Проводимость — способность проводить возбуждение.
Сократимость — способность мышцы изменять свою длину или напряжение в ответ на действие раздражителя.
Лабильность – по Н.Е.Введенскому, наибольшее число потенциалов действия, которое возбудимая ткань способна воспроизвести в единицу времени (1 сек.) под влиянием частых приложений к ней раздражений (лабильность мышечного волокна равна 20-30 импульсов в секунду, нервного около 1000).
§ Автоматия – способность генерировать импульсы без внешнего раздражения (свойство характерно для сердечной мышцы и для гладких мышц).
Скелетные (поперечнополосатые) мышцы в организме играют роль своеобразных «машин», преобразующих химическую энергию непосредственно в механическую и тепловую. Сокращение мышц возникает в ответ на электрические импульсы, приходящие к ним от альфа-мотонейронов – нервных клеток, лежащих в передних рогах спинного мозга.
Мышцы и иннервирующие их мотонейроны составляют нервно-мышечный аппарат человека.
Организм человека обладает выраженной способностью адаптироваться к постоянно меняющимся условиям внешней среды. В основе приспособительных реакций организма лежит универсальное свойство живой ткани — раздражимость — способность отвечать на действие раздражающих факторов изменением структурных и функциональных свойств. Раздражимостью обладают все ткани животных и растительных организмов. В процессе эволюции происходила постепенная дифференциация тканей, осуществляющих приспособительную деятельность организма. Раздражимость этих тканей достигла наивысшего развития и трансформировалась в новое свойство — возбудимость. Под этим термином понимают способность ткани отвечать на раздражение специализированной реакцией — возбуждением. Возбуждение — это сложный биологический процесс, который характеризуется специфическим изменением процессов обмена веществ, теплообразования, временной деполяризацией мембраны клеток и проявляющийся специализированной реакцией ткани (сокращение мышцы, отделение секрета железой и т. д.).
Возбудимостью обладают нервная, мышечная и секреторная ткани, их объединяют в понятие "возбудимые ткани".
Возбудимость различных тканей неодинакова. Мерой возбудимости является порог раздражения — минимальная сила раздражителя, которая способна вызвать возбуждение. Менее сильные раздражители называются подпороговыми, а более сильные — сверхпороговыми. Раздражителем живой клетки может быть любое изменение внешней или внутренней среды, если оно достаточно велико, возникло достаточно быстро и продолжается достаточно долго.
Природа возбуждения
Первые попытки последовательной разработки учения о "животном электричестве" связаны с именем Л. Гальвани. Э. Дюбуа-Реймон впервые показал, что наружная поверхность мышцы заряжена положительно по отношению к ее внутреннему содержимому. Следовательно, в состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая затем была названа мембранным потенциалом покоя или мембранным потенциалом. Его величина у разных клеток колеблется от 60 до 90 мВ.
А. Ходжкин, А. Хаксли и Б. Катц в 50-х годах 20 века объяснили причины возникновения мембранного потенциала покоя, для чего они существенно переработали ранее существовавшие представления и создали мембранно-ионную теорию. Согласно их взглядам мембранный потенциал покоя (МПП) обусловлен неодинаковой концентрацией ионов натрия, калия, кальция, хлора внутри клетки и во внеклеточной жидкости, а также неодинаковой проницаемостью для этих ионов поверхностной мембраны клетки (рис. 2.4). Цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Следовательно, в состоянии покоя существует асимметрия концентрации ионов внутри клетки и в окружающей ее среде.

Рис. 2.4. Регистрация потенциала покоя
В мембране имеются ионные каналы, образованные макромолекулами белка, пронизывающих липидный слой. Каналы мембраны делятся на неспецифические (каналы утечки) и специфические (селективные, обладающие способностью пропускать только определенные ионы). Неспецифические каналы пропускают различные ионы и открыты постоянно. Специфические каналы открываются и закрываются в ответ на изменения МПП.
шиза. 1. Общие свойства возбудимых тканей. Возбудимость
Эти каналы называются потенциалозависимыми.
В состоянии физиологического покоя мембрана нервных волокон в 25 раз лучше проницаема для K+ , чем для Na+ .
Выход положительно заряженных ионов калия приводит к появлению положительного заряда на наружной поверхности мембраны. Органические анионы — крупномолекулярные соединения, которые несут отрицательный заряд, и для которых мембрана клетки непроницаема, придают в этих условиях внутренней поверхности мембраны отрицательный заряд (рис.2.5).

Рис.2.5. Концентрация основных ионов внутри и вне клетки.
В состоянии покоя существуют небольшие перемещения K+ и Na+ через мембрану по их концентрационному градиенту (табл.2.2), K+ больше, чем Na+ .
Табл. 2.2.
©2015-2018 poisk-ru.ru
Все права принадлежать их авторам. Данный сайт не претендует на авторства, а предоставляет бесплатное использование.
Нарушение авторских прав и Нарушение персональных данных
II. Понятие о раздражимости, возбудимости и возбуждении. Классификация раздражителей
Раздражимость – это способность клеток, тканей, организма в целом переходить под воздействием факторов внешней или внутренней среды из состояния физиологического покоя в состояние активности. Состояние активности проявляется изменением физиологических параметров клетки, ткани, организма, например, изменением метаболизма.
Возбудимость – это способность живой ткани отвечать на раздражение активной специфической реакцией – возбуждением, т.е. генерацией нервного импульса, сокращением, секрецией. Т.о., возбудимость характеризует специализированные ткани – нервную, мышечные, железистые, которые называются возбудимыми.
Возбуждение– это комплекс процессов реагирования возбудимой ткани на действие раздражителя, проявляющийся изменением мембранного потенциала, метаболизма и т.д. Возбудимые ткани обладают проводимостью. Это способность ткани проводить возбуждение. Наибольшей проводимостью обладают нервы и скелетные мышцы.
Раздражитель – это фактор внешней или внутренней среды действующий на живую ткань.
Процесс воздействия раздражителя на клетку, ткань, организм называется раздражением.
Все раздражители делятся на следующие группы:
1. По природе : а) физические (электричество, свет, звук, механические воздействия и т.д.); б) химические (кислоты, щелочи, гормоны и т.д.); в) физико-химические (осмотическое давление, парциальное давление газов и т.д.); г) биологические (пища для животного, особь другого пола); д) социальные (слово для человека).
2. По месту воздействия : а) внешние (экзогенные); б) внутренние (эндогенные).
3. По силе : а) подпороговые; б) пороговые (раздражители минимальной, силы, при которой возникает возбуждение); в) сверхпороговые (силой выше пороговой).
4. По физиологическому характеру : а) адекватные (физиологичные для данной клетки или рецептора, которые, приспособились к нему в процессе эволюции, например, свет для фоторецепторов глаза); б) неадекватные.
5. Если реакция на раздражитель является рефлекторной , то выделяют также:
а) безусловно-рефлекторные раздражители; б) условно-рефлекторные.
ІІІ. Потенциал покоя (МПП)
Потенциал покоя - относительно стабильная разность электрических потенциалов между наружной и внутренней сторонами клеточ-ной мембраны. Его величина обычно варьи-рует в пределах 30-90 мВ (в волокнах ске-летной мышцы - 60-90 мВ, в нервных клет-ках - 50-80 мВ, в гладких мышцах - 30- 70 мВ, в сердечной мышце - 80-90 мВ).
ПП играет исключительно важную роль в жизнедеятельности самой клетки и организ-ма в целом, поскольку является основой для возникновения возбуждения (потенциала действия), с помощью которого нервная сис-тема воспринимает и перерабатывает инфор-мацию, регулирует деятельность внутренних органов и опорно-двигательного аппарата посредством запуска процессов возбуждения и сокращения в мышце. Нарушение процес-сов возбуждения в кардиомиоцитах ведет к остановке сердца.
Согласно мембранно-ионной теории (Бернштейн, Ходжкин, Хаксли, Катц, 1902- 1952), непосредственной причиной формиро-вания ПП является неодинаковая концентра-ция анионов и катионов внутри и вне клетки.
Различные ионы распреде-лены неравномерно по обе стороны клеточ-ной мембраны, во-первых, вследствие неоди-наковой проницаемости клеточной мембра-ны для различных ионов, во-вторых - в ре-зультате работы ионных насосов, транспор-тирующих ионы в клетку и из клетки вопре-ки концентрационному и электрическому градиентам.
Роль проницаемости клеточной мембраны в формировании ПП . Проницаемость клеточной мембраны - это ее способность пропускать воду, незаряженные и заряженные частицы (ионы) согласно законам диффузии и фильт-рации. Термин «проводимость» следует использо-вать только лишь применительно к заряжен-ным частицам. Следовательно, проводи-мость - это способность заряженных частиц (ионов) проходить через клеточную мембра-ну согласно электрохимическому градиенту.
Na+ и К+ в покоящейся клетке перемещаются через мембрану согласно законам диффузии, при этом К+ из клетки выходит в значитель-но большем количестве, чем входит Na+ в клетку, поскольку проницаемость клеточной мембраны для К+ примерно в 25 раз больше проницаемости для Na+.
Органические анионы из-за своих больших размеров не могут выходить из клетки, поэ-тому внутри клетки в состоянии покоя отри-цательных ионов оказывается больше, чем положительных. По этой причине клетка из-нутри имеет отрицательный заряд. Интерес-но, что во всех точках клетки отрицательный заряд практически одинаков. Об этом свиде-тельствует одинаковая величина ПП при вве-дении микроэлектрода на разную глубину внутрь клетки, как это имело место в опытах Ходжкина, Хаксли и Катца. Гигантский аксон кальмара (его диаметр около 1 мм) в этом опыте находился в морской воде, один электрод вводился в аксон, другой помещали в морскую воду. Заряд внутри клетки являет-ся отрицательным как абсолютно (в гиалоплазме клетки содержится больше анионов, нежели катионов), так и относительно на-ружной поверхности клеточной мембраны. Однако превышение абсолютного числа анионов над числом катионов в клетке чрез-вычайно мало. Но этого различия достаточно для создания разности электрических потен-циалов внутри и вне клетки.
Главным ионом, обеспечивающим форми-рование ПП, является ион К+. Об этом сви-детельствуют результаты опыта с перфузией внутреннего содержимого гигантского аксо-на кальмара солевыми растворами. При уменьшении концентрации К+ в перфузате ПП уменьшается, при увеличении концент-рации К+ ПП увеличивается. В покоящейся клетке устанавливается динамическое равно-весие между числом выходящих из клетки и входящих в клетку ионов К+. Электрический и концентрационный градиенты противодей-ствуют друг другу: согласно концентрацион-ному градиенту К+ стремится выйти из клет-ки, отрицательный заряд внутри клетки и по-ложительный заряд наружной поверхности клеточной мембраны препятствуют этому. Когда концентрационный и электрический градиенты уравновесятся, число выходящих из клетки ионов К+ сравнивается с числом входящих ионов К+ в клетку. В этом случае на клеточной мембране устанавливается так называемый равновесный калиевый потен-циал. Равновесный потенциал для любого иона можно рассчитать по формуле Нернста, а для нескольких по формуле Гольдмана-Ходжкина-Катца
В целом ПП - это производное равновесных потенциалов всех ионов, находя-щихся внутри и вне клетки и поверхностных зарядов клеточной мембраны.
Вклад Na+ и Сl- в создание ПП. Проницае-мость клеточной мембраны в покое для Na+ очень низкая - намного ниже, чем для К+, тем не менее она имеет место, поэтому ионы Na+, согласно концентрационному и элект-рическому градиентам, стремятся и в неболь-шом количестве проходят внутрь клетки. Это ведет к уменьшению ПП, так как на внешней поверхности клеточной мембраны суммарное число положительно заряженных ионов уменьшается, хотя и незначительно, а часть отрицательных ионов внутри клетки нейтра-лизуется входящими в клетку положительно заряженными ионами Na+. Вход Na+ внутрь клетки уменьшает ПП. Что касается Сl— , его влияние на величину ПП противоположно влиянию Na+ и зависит от проницаемости клеточной мембраны для Сl— (она в 2 раза ниже, чем для К+). Дело в том, что Сl— , со-гласно концентрационному градиенту, стремится и проходит в клетку. Концентрации ионов К+ и Сl— близки между собой. Но Сl— находится в основном вне клетки, а К+ - внутри клетки. Препятствует входу Сl— в клетку электрический градиент, поскольку заряд внутри клетки отрицательный, как и заряд Сl— . Наступает равновесие сил кон-центрационного градиента, способствующего входу Сl— в клетку, и электрического гради-ента, препятствующего входу Сl— в клетку. Поэтому внутриклеточная концентрация Сl— равна всего лишь 5-10 ммоль/л, а вне клет-ки - 120-130 ммоль/л. При поступлении Сl— внутрь клетки число отрицательных зарядов вне клетки несколько уменьшается, а внутри клетки увеличивается: Сl— добавляется к крупным белковой природы анионам, нахо-дящимся внутри клетки. Эти анионы из-за своих больших размеров не могут пройти через каналы клеточной мембраны нару-жу клетки - в интерстиций. Таким образом, Сl-, проникая внутрь клетки, увеличивает ПП. Частично, как и вне клетки, Na+ и Сl— внутри клетки нейтрализуют друг друга. Вследствие этого совместное поступление Na+ и Сl— внутрь клетки не сказывается су-щественно на величине ПП.
Роль поверхностных зарядов клеточной мембраны и ионов Са2+ в формировании ПП. Наружная и внутренняя поверхности клеточ-ной мембраны несут собственные электри-ческие заряды, преимущественно с отрица-тельным знаком. Это полярные молекулы клеточной мембраны - гликолипиды, фосфолипиды, гликопротеиды. Фиксированные наружные отрицательные заряды, нейтрали-зуя положительные заряды внешней поверх-ности мембраны, уменьшают ПП. Фиксиро-ванные внутренние отрицательные заряды клеточной мембраны, напротив, суммируясь с анионами внутри клетки, увеличивают ПП. Роль ионов Са2+ в формировании ПП заклю-чается в том, что они взаимодействуют с на-ружными отрицательными фиксированными зарядами мембраны клетки и отрицательны-ми карбоксильными группами интерстиция и нейтрализуют их, что ведет к увеличению и стабилизации ПП.
Таким образом, ПП - это алгебраическая сумма не только всех зарядов ионов вне и внут-ри клетки, но также алгебраическая сумма отрицательных внешних и внутренних поверх-ностных зарядов самой мембраны.
Роль ионных насосов в формировании ПП . В результате непрерывного перемещения различных ионов через клеточную мембрану их концентрация внутри и вне клетки посте-пенно должна выравниваться. Однако, не-смотря на постоянную диффузию ионов (утечку ионов), ПП клеток остается на одном уровне. Следовательно, кроме собственных ионных механизмов формирования ПП, свя-занных с различной проницаемостью клеточ-ной мембраны, имеется активный механизм поддержания градиентов концентрации раз-личных ионов внутри и вне клетки. Им явля-ются ионные насосы, в частности Na/K-насос (помпа). В результате сопря-женного транспорта Na+ и К+ поддерживается постоянная разность концентраций этих ионов внутри и вне клетки. Одна молекула АТФ обеспечивает один цикл работы Na/K-насоса - перенос трех ионов Na+ за пределы клетки и двух ионов К+ внутрь клетки.
Возбудимость и проводимость - свойства, характерные для ткани
Асим-метричный перенос ионов Na/K-насосом поддерживает избыток положительно заря-женных частиц на наружной поверхности клеточной мембраны и отрицательных заря-дов внутри клетки, что позволяет считать Na/K-насос структурой электрогенной, до-полнительно увеличивающей ПП примерно на 5-10 мВ (в среднем около 10 % у разных возбудимых клеток - у одних больше, у дру-гих меньше). Данный факт свидетельствует о том, что решающим фактором в формирова-нии ПП является селективная проницае-мость клеточной мембраны для разных ионов. Если уравнять проницаемость клеточ-ной мембраны для всех ионов, то ПП будет составлять только 5-10 мВ - за счет работы N/K-помпы.
Нормальная величина ПП является необ-ходимым условием возникновения процесса возбуждения клетки, т.е. возникновения и распространения потенциала действия, ини-циирующего специфическую деятельность клетки.
ІІІ. Электротонический и локальный потенциалы (рис.6)
Если воздействовать на клетку раздражителем в размере 1-50% от порогового, клетка ответит электротоничсеким потенциалом – сдвигом МП клетки. Это пассивная реакция клетки на электрический раздражитель; состояние ионных каналов и транспорт ионов не изменяется, либо изменяется совсем незначительно на доли миллисекунд. ЭП не является физиологической реакцией клетки и т.о. не является возбуждением.
Если же на клетку подействовать подпороговым током (50-99% от пороговой величины) – развивается пролонгированный сдвиг МП – локальный ответ. Это активная реакция клетки на раздражитель, однако, состояние ионных и транспорт ионов меняется незначительно. ЛО называют местным возбуждением, т.к. оно не распространяется по мембранам возбудимых клеток, также это не способная к распространению деполяризация мембраны. В основном она обусловлена перемещением ионов Na + в клетку. Вследствие этого уровень поляризации мембраны уменьшается.
Свойства ЛО:
- распространяется с затуханием
- подчиняется закону градуальности (постепенное нарастание или затухание)
- может суммироваться
- нет периода рефрактерности
- имеет фазу деполяризации и реполяризации
 рис. 6
рис. 6
Предыдущая12345678910111213141516Следующая
Возбудимые ткани — это ткани, котоpые способны воспpинимать действие pаздpажителя и отвечать на него пеpеходом в состояние возбуждения
К возбудимым тканям относятся тpи вида тканей — это неpвная, мышечная и железистая
Возбудимые ткани обладают pядом общих и частных свойств.
Общими свойствами возбудимых тканей являются:
1.Раздpажимость
2.Возбудимость
Пpоводимость
Раздpажимость — это способность клетки, ткани или оpгана воспpинимать действие pаздpажителя изменением метаболизма, стpуктуpы и функций
Раздpажимость является унивеpсальным свойством всего живого и является основой пpиспособительных pеакций живого оpганизма к постоянно меняющимся условиям внешней и внутpенней сpеды.
Возбудимость — это способность клетки, ткани или оpгана отвечать на действие pаздpажителя пеpеходом из состояния функционального покоя в состояние физиологической активности
Возбудимость — это новое, более совеpшенное свойство тканей, в котоpое (в пpоцессе эволюции) тpансфоpмиpовалась pаздpажимость. Разные ткани обладают pазличной возбудимостью: неpвная > мышечная > железистая
Меpой возбудимость является поpог pаздpажения
Поpог pаздpажения — это минимальная сила pаздpажителя, способная вызвать pаспpостpоняющееся возбуждение
Возбудимость и поpог pаздpажения находятся в обpатной зависимости (чем > возбудимость, тем < поpог pаздpажения)
Возбудимость зависит от:
1.Величины потенциала покоя
2.Уpовня кpитической деполяpизации
Потенциал покоя — это pазность потенциалов между внутpенней и наpужней повеpхностями мембpаны в состояни покоя
Уpовень кpитической деполяpизации — это та величина мембpанного потенциала, котоpую необходимо достичь, чтобы возбуждение носило pаспpостpаняющийся хаpактеp
Разница между значениями потенциала покоя и уpовнем кpитической деполяpизации опpеделяет поpог деполяpизации (чем < поpог деполяpизации, тем > возбудимость)
Пpоводимость — это способность пpоводить возбуждение
Пpоводимость опpеделяется:
1.Стpоением ткани
2.Функциональными особенностями ткани
Возбудимостью
Память — это способность фиксиpовать изменения функционального состояния клетки, ткани, оpгана и оpганизма на молекуляpном уpовне
Опpеделяется генетической пpогpаммой
Позволяет отвечать на действие отдельных, значимых для оpганизма pаздpажителей с опеpежением
К частным свойствам возбудимых тканей относятся:
1.Сокpатимость
2.Секpетоpная деятельность
Автоматия
Сокpатимость — способность мышечных стpуктуp изменять длину или напpяжение в ответ на возбуждение
Зависит от вида мышечной ткани
Секpетоpная активность — это способность выделять медиатоp или секpет в ответ на возбуждение
Теpминали нейpонов секpетиpуют медиатоpы
Железистые клетки экскpетиpуют пот, слюну, желудочный и кишечный сок, желчь, а также инкpетиpуют гоpмоны и биологически активные вещества
Автоматия — это способность самостоятельно возбуждаться, то есть возбуждаться без действия pаздpажителя или пpиходящего неpвного импульса
Хаpактеpна для сеpдечной мышцы, гладкой мускулатуpы, отдельных неpвных клеток центpальной неpвной системы
Для возбудимых тканей хаpактеpно 2 вида функциональной активности
Физиологический покой — состояние без пpоявлений специфической деятельности (пpи отсутствии действия pаздpажителя)
Возбуждение — активное состояние, котоpое пpоявляется стpуктуpными и физико-химическими сдвигами (специфическая фоpма pеагиpования в ответ на действие pаздpажителя или пpиходящего неpвного импульса)
Различные виды функциональной активности опpеделяются стpуктуpой, свойством и состоянием плазматических мембpан
№9 Функции: 1. Барьерная – мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии.
2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»).
3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах).
4.Высвобождение нейромедиаторов в синаптических окончаниях.
Жидкостно-мозаичная модель Сингера и Николсона:
В фосфолипидном бислое интегрированы глобулярные белки, полярные участки которых образуют гидрофильную поверхность в водной фазе. Эти интегрированные белки выполняют различные функции, в том числе рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул.
Некоторые белковые молекулы свободно диффундируют в плоскости липидного слоя; в обычном состоянии части белковых молекул, выходящие по разные стороны клеточной мембраны, не изменяют своего положения.
Особая морфология клеточных мембран определяет их электрические характеристики, среди которых наиболее важными являются емкость и проводимость.
Емкостные свойства в основном определяются фосфолипидным бислоем, который непроницаем для гидратированных ионов и в то же время достаточно тонок (около 5 нм), чтобы обеспечивать эффективное разделение и накопление зарядов и электростатическое взаимодействие катионов и анионов. Активный транспорт — транспорт веществ через мембрану, который осуществляется против градиента концентрации и требует значительных затрат энергии . Одна треть основного обмена тратиться на активный транспорт.
Активный транспорт бывает:
1. Первично-активый — такой транспорт, для обеспечения которого используется энергия макроэргов — АТФ, ГТФ, креатинфосфат. Например : Калиево-натриевый насос — важная роль в процессах возбудимости в клетке. Он вмонтирован в мембрану.
Калиево-натриевый насос — фермент калий-натриевая АТФаза.
Возбудимые ткани и их основные свойства
Этот фермент — белок. Он существует в мембране в виде 2-х форм:
Е 1, Е 2
В ферментах существует активный участок , который взаимодействует с калием и с натрием. Когда фермент находится в форме Е 1 , его активный участок обращен внутрь клетки и обладает высоким сродством к натрию , а значит способствует его присоединению (3 атома Na). Как только натрий присоединяется, происходит конформация этого белка, которая перемещает 3 атома натрия через мембрану и с наружной поверхности мембраны натрий отсоединяется. При этом происходит переход фермента из формы Е 1 в Е 2. Е 2 имеет активный участок, обращенный к наружной поверхности клетки, обладает высоким сродством к калию . При этом 2 атома К присоединяется к активному участку фермента, изменяется конформация белка и калий перемещается внутрь клетки. Это происходит с большой затратой энергии , так как фермент АТФаза постоянно расщепляет энергию АТФ.
2. Вторично-активный — это транспорт, который осуществляется тоже против градиента концентрации, но на это перемещение тратится не энергия макроэргов, а энергия электрохимических процессов, которая возникает при движении каких-либо веществ через мембрану при первично-активном транспорте.
Например : Сопряженный транспорт натрия и глюкозы, энергия — за счет перемещения натрия в калиево-натриевом насосе.
Классическим примером вторично-активного транспорта выступает натрий – Н (аш)-обменник — когда обмениваются натрий и водород (это тоже вторично-активный транспорт).
Способы транспортировки через мембрану:
1. Унипорт — это такой вид транспорта веществ через мембрану, когда переносчиком или каналом транспортируется одно вещество (Na-каналы)
2. Симпорт — это такой вид транспорта, когда 2 или более веществ в своем транспорте через мембрану взаимосвязаны и транспортируются вместе в одном направлении. (Na и глюкоза — в клетку) Это вид сопряженного транспорта
3. Антипорт — такой сопряжѐнный вид транспорта, когда его участники друг без друга не могут транспортироваться, но потоки идут навстречу друг другу (К-Na-насос-активный вид транспорта).
Эндоцитоз, экзоцитоз — как формы транспорта веществ через мембрану.
Дата публикования: 2015-03-29; Прочитано: 11717 | Нарушение авторского права страницы
studopedia.org — Студопедия.Орг — 2014-2018 год.(0.003 с)…
Что такое возбуждение, какие ткани относятся к возбудимым?
Возбуждение
Какой феномен отражает состояние возбуждения мышечной клетки?
Заряд клеточной мембраны
Что такое возбудимость?
Способность к возбуждению.
5.Как можно оценить возбудимость различных клеток, приведите пример?
По параметру пороговой силы. Чем меньше пороговая сила,тем больше возбудимость. Самая возбудимая ткань – нервная.
У клетки А КУД 60 мв, мембранный потенциал 80 мв, у клетки В КУД 60 мв, мембранный потенциал 90 мв, какая клетка является более возбудимой, почему?
Клетка А обладает большей возбудимостью, так как возбуждение оценивает по пороговой силе раздражителя(его минимальная сила, при которой клетка возбуждается).
У клетки А КУД 60 мв, мембранный потенциал(МП) 80 мв, у клетки В КУД 70 мв, МП 90 мв, какая клетка является более возбудимой, ответ докажите?
Клетки обладают одинаковой возбудимостью, так как возбуждение оценивает по пороговой силе раздражителя(его минимальная сила, при которой клетка возбуждается), а они у клеток одинаковы.
8.Какие электрофизиологические характеристики клеточной мембраны предопределяют возбудимость клеток? Приведите пример.
Мембранный потенциал и КУД.
9.Приведите пример реагирования возбудимой ткани на пороговые и сверхпороговые раздражители по закону «силовых соотношений». Объясните причину такого реагирования.
Возбудимая ткань на пороговые и сверхпороговые раздражители реагирует по принципу чем больше ток, тем больше ответная реакция. Отдельные клетки в силу разных причин обладают разным порогом возбудимости, поэтому в начале в мышце сокращаются наиболее возбудимые клетки (пороговое сокращение), а затем по мере увеличения силы раздражителя в сокращение вовлекаются все новые и новые менее возбудимые клетки. Когда все клетки возбуждены, увеличение амплитуды раздражителя не вызывает прироста ответной реакции (сокращения
10.Приведите пример реагирования возбудимой ткани по закону «все или ничего». Объясните причину такого реагирования.
На пороговые и сверхпороговые раздражители ткань реагируют сокращением одинаковой силы. Это характерно для однородных систем (1 клетка, также сердечная. Мышца сердца представляет собой так называемый функциональный синцитий, а скелетная мышца – симпласт.
Что отражает МПП нейрона, чему он равен, как можно определить его величину.
Разность потенциалов между наружной и внутренней поверхностями мембраны клетки в покое.
Охарактеризуйте ионные механизмы, обеспечивающие возникновение мембранного потенциала в нервных и мышечных клетках?
Калиевый механизм – основной механизм, который лежит в основе поляризации клеточной мембраны. Также играет роль K – Na насос и ток Na из клетки.
13.Как изменится МПП при повышении внеклеточной концентрации ионов К+, как это отразится на возбудимости клетки?
МПП снизится, так как снизится разность зарядов.
Возбудимость и проводимость свойства характерные для ткани
Возбудимость увеличится вследствие изменения МПП.
Как изменится МПД после обработки клетки блокатором потенциалуправляемых натриевых каналов мембраны?
снизится
Решите задачу – внутриклеточный потенциал мышечной клетки равен -80 мВ. Чему равен мембранный потенциал покоя?
16.Расшифруйте понятия – поляризация, деполяризация, реполяри-зация, гиперполяризация.
Поляризация – ассиметричное распределение зарядов относительно клеточной мембраны.
Деполяризация – уменьшение напряжения на мембране.
Гиперполяризация – увеличение напряжения на мембране.
Реполяризация – восстановление мембранного потенциала после де- и гиперполяризации.
Нарисуйте электрограмму возникновения МПД при пороговом и сверпорпоговом раздражении мышечной клетки.

- локальный ответ (пороговая деполяризация)
- фаза быстрой деполяризации
- перескок (овершут) – фаза перезарядки
- фаза реполяризации
- следовой отрицательный потенциал
- следовой положительный потенциал
Объясните ионный механизм возникновения МПД.
Идет открытие управляемых натриевых каналов под влиянием концентрационного и электрического градинетов. Число управляемых натриевых каналов больше неуправляемых калиевых. В результате происходит перезарядка мембраны. В фазу перескока ток К из клетку начинает преобладать над током Na в клетку и заряд начинает падать.
Что такое полезное время, что такое хронаксия.
Наименьшая длительность раздражения силой в одну реобазу необходимая для возникновения возбуждения называется полезным временем . Реобаза – та минимальная сила раздражение, на которую при практически неограниченном большой длительностью ее действия возникнет минимальная ответная реакция.
Хронаксия – минимальное время в течении которого раздражитель с силой равной двум реобазам вызывает возбуждение
Что такое «гарантийный фактор» проведения возбуждения?
Фактор надежности (гарантийный фактор) = ПД: возбудимость нервного волокна.в норме 5-6 единиц
Что такое возбуждение, какие ткани относятся к возбудимым?
Возбуждение – это процесс генерации потенциала действия под воздействием пороговых и сверхпороговых раздражителей. Возбудимые ткани: мышечная, нервная и железистая.
2.Какую функцию выполняет возбуждение. Приведите примеры.
Возбуждение в возбудимых тканях запускает специальные реакции. Мышцы – сокращение, нервы – импульс, выделение медиатора, железа – секреция.
Раздражимость
Раздражимостью называют способность живых систем под влиянием раздражителей переходить из состояния физиологического покоя в состояние активности. Формы проявления раздражимости весьма разнообразны. У одних клеток при этом изменяются форма, структура, их рост и процесс деления; в других образуются различные химические соединения, совершается та или иная работа, движение.
Раздражителем живой клетки или организма как целого может оказаться любое изменение внешней среды или внутреннего состояния организма, если оно достигает определенной величины.
Среди раздражителей различают физические (температура, укол, давление, свет, звук, электрические раздражители), физико-химические (изменения осмотического давления, активной реакции среды, электролитного состава, коллоидного состояния) и химические (химические пищи, химические соединения, образующиеся в организме,- гормоны, продукты обмена веществ и т. п.).
Рис. 17. Первый (I ) и второй (II) опыты
Естественными раздражителями клеток, вызывающими их деятельность, являются нервные импульсы.
Возбудимость
Клетки нервной и мышечной тканей приспособлены к осуществлению быстрых реакций на раздражение. Клетки этих тканей называют возбудимыми, а их способность отвечать на раздражение возбуждением называют возбудимостью.
Возбуждение способно перемещаться из одного места клетки в другое, из одной клетки в другую.
 Рис. 18.
Последовательные этапы приготовления нервно-мышечного препарата.
Рис. 18.
Последовательные этапы приготовления нервно-мышечного препарата.
Возбуждение характеризуется комплексом химических, функциональных, физико-химических, электрических явлений. Обязательным признаком возбуждения является изменение электрического состояния поверхностной клеточной мембраны. Именно электрические явления обеспечивают проведение возбуждения в возбудимых тканях.
Биоэлектрические явления
Возникновение и распространение возбуждения связано с изменением электрического заряда живой ткани, с так называемыми биоэлектрическими явлениями.
Электрические явления у животных известны давно. Еще в 1776 г. они были описаны у электрического ската. Началом же экспериментального изучения электрических явлений в животных тканях следует считать опыты итальянского врача Луиджи (1791). В опытах он использовал препараты задних конечностей лягушки, соединенных с позвоночником. Подвешивая эти препараты на медном крючке к железным перилам балкона, он обратил внимание, что, когда конечности лягушки раскачивались ветром, их мышцы сокращались при каждом прикосновении к перилам. На основании этого пришел к выводу, что подергивания лапок были вызваны «животным электричеством», зарождающимся в спинном мозге лягушки и передаваемым по металлическим проводникам (крючку и перилам балкона) к мышцам препарата.
 Рис. 19.
Схема вторичного сокращения.
Рис. 19.
Схема вторичного сокращения.
Против этого положения Гальвани о «животном электричестве» выступил физик Александр . В 1792 г. повторил опыты Гальвани и установил, что описанные Гальвани явления нельзя считать «животным электричеством». В опыте Гальвани источником тока служил не спинной мозг лягушки, а цепь, образованная из разнородных металлов - меди и железа.
Был прав. Первый опыт Гальвани не доказывал наличия «животного электричества», но эти исследования привлекли внимание ученых к изучению электрических явлений в живых образованиях.
В ответ на возражение Вольта Гальвани произвел второй опыт, уже без участия металлов. При этом также наблюдалось сокращение лапки лягушки.
Опыт Гальвани. Для этого обездвижьте лягушку и перережьте ее поперек в области верхних грудных позвонков. Захватив остаток позвоночника салфеточкой, снимите с задних конечностей кожу, а затем пинцетом удалите остатки внутренностей. Становятся хорошо видны нервные стволики крестцового сплетения, расположенные с обеих сторон позвоночника пучками. Подведите под оба пучка нервных волокон одну пластинку пинцета Гальвани, а другой пластинкой пинцета прикоснитесь к нервам сверху. Мышцы лапок при этом сокращаются (рис. 17, I). Пинцет Гальвани состоит из цинковой и медной пластинок. Объясните, почему сокращаются мышцы лапок в опыте Гальвани.
 Рис. 20.
Рис. 20.
А - одноканальный; Б - двухканальный; справа - кончик электрода около тела нейрона
А теперь приготовьте нервно-мышечный препарат лягушки.
Основные этапы приготовления нервно-мышечного препарата приведены на рисунке 18.
Лягушку обездвижьте. Возьмите ее левой рукой за бедра (в этом положении хорошо выделяется позвоночник) и перережьте позвоночник на 1-1,5 см выше места отхождения тазовых костей (рис. 18, 1 ). Свисающую переднюю часть туловища и внутренности удалите. Остаток позвоночника крепко держите пинцетом или левой рукой. Другим пинцетом захватите кожу около позвоночника и тяните ее вниз, чтобы, выворачивая, снять с конечностей (рис. 18, 2 ). Конечности положите на чистую тарелку и залейте раствором Рингера. Руки вымойте или тщательно вытрите от слизи, покрывающей кожу лягушки. Захватите пинцетом или рукой кусочек позвоночника и подогните его вниз так, чтобы конечности висели под углом к позвоночнику и хорошо выделялась копчиковая кость (рис. 18,3).
Осторожно вырежьте копчиковую кость. Ножницы при этом держите как можно ближе к кости, чтобы не повредить идущие параллельно с обеих сторон нервы. Вырезав копчик, положите препарат на тарелку и разделите его на две половины. Для этого перережьте вдоль сначала остаток позвоночника, а затем лобковое сочленение (рис. 18, 4).
 Рис. 21.
А
с помощью микроэлектрода:
Рис. 21.
А
с помощью микроэлектрода:
М - микроэлектрод; И - индифферентный электрод. Луч на экране осциллографа Б показывает, что до прокола микроэлектро дом мембраны разность потенциалов меж ду М и И была равна нулю. В момент прокола (показан стрелкой) обнаружена разность потенциалов, указывающая, что внутренняя сторона мембраны заряжена электроотрицательно по отношению к ее наружной поверхности.
Одну конечность оставьте как запасную, сохраняя ее в растворе Рингера; другую положите на спинную сторону и отделите ножницами подвздошную кость. Захватив пинцетом кусочек позвоночника, отведите в сторону седалищный нерв и удалите подвздошную кость. При помощи двух пинцетов раздвиньте мышцу на спинной поверхности бедра по средней линии (рис. 18, 5). Осторожно не касаясь ножницами и пинцетом нерва, отделите его от окружающих тканей, вдоль бедра до колен. (Лучше это делать стеклянным крючком.) Нерв отведите в сторону и освободите бедренную кость от мышц (рис. 18, б). На голени отделите от кости икроножную мышцу, подрезав ахиллово сухожилие, и привяжите к нему нитку. Голень и стопу отрежьте ниже колена (рис. 18, 7). Препарат положите в стакан с раствором Рингера.
Проделайте второй опыт Гальвани (сокращение без металла). Для этого нервно-мышечный препарат положите на дощечку. Отрежьте кусочек мышцы и стеклянным крючком быстро набросьте нерв препарата на пораненный участок мышцы так, чтобы он одновременно коснулся поврежденной и неповрежденной поверхности мышцы (рис. 17, II). Мышца при этом сокращается. Объясните, почему это происходит.
Гальвани все же оказался прав в своем утверждении о существовании «животного электричества», что позже было подтверждено исследованиями других ученых.
В этом отношении интересны опыты Маттеучю, получившие название вторичного сокращения.
На мышцу одного нервно-мышечного препарата набросьте нерв другого нервно-мышечного препарата (рис. 19) и раздражайте электрическим током нерв первого препарата. Вы наблюдаете сокращение мышцы и второго препарата. Это объясняется тем, что при возбуждении в мышце первого препарата возникают токи действия, которые вызывают возбуждение второго нервно-мышечного препарата.
В дальнейшем в изучение биоэлектрических явлений очень важный вклад внесли русские ученые, среди них И. М. Сеченов, обнаруживший с помощью гальванометра электрические явления в головном мозге, Н. Е. Введенский, А. Ф. Самойлов и др.
В настоящее время имеются весьма совершенные, высокочувствительные приборы (электронно-лучевые трубки с электронными усилителями), позволяющие регистрировать электрические явления в тканях и органах.
Потенциал покоя и потенциал действия мышц
Между наружной поверхностью клетки и ее цитоплазмой в состоянии покоя существует разность потенциалов около 60-90 мВ, причем поверхность клетки заряжена электроположительно по отношению к цитоплазме. Эту разность потенциалов называют потенциалом покоя или мембранным потенциалом. Точное измерение потенциала покоя возможно только с помощью микроэлектродов, предназначенных для внутриклеточного отведения токов, очень мощных усилителей и чувствительных регистрирующих приборов - осциллографов.
 Рис. 22.
Потенциал действия аксона кальмара, отводимый с помощью внутриклеточного электрода.
Рис. 22.
Потенциал действия аксона кальмара, отводимый с помощью внутриклеточного электрода.
Микроэлектрод (рис. 20) представляет собой тонкий стеклянный капилляр, кончик которого имеет диаметр около микрона. Этот капилляр заполняют солевым раствором, погружают в него металлический электрод и соединяют с усилителем и осциллографом (рис. 21). Как только микроэлектрод прокалывает покрывающую клетку мембрану, луч осциллографа отклоняется вниз из своего исходного положения и устанавливается на новом уровне. Это свидетельствует о наличии разности потенциалов между наружной и внутренней поверхностью клеточной мембраны.
Наиболее полно происхождение потенциала покоя объясняет так называемая мембран но-ионная теория. Согласно этой теории все клетки покрыты мембраной, имеющей неодинаковую проницаемость для различных ионов. В связи с этим внутри клетки в цитоплазме в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем на поверхности клетки. В состоянии покоя клеточная мембрана более проницаема для ионов калия, чем для ионов натрия.
Диффузия положительно заряженных ионов калия из цитоплазмы на поверхность клетки придает наружной поверхности мембраны положительный заряд.
Таким образом, поверхность клетки в покое несет на себе положительный заряд, тогда как внутренняя сторона мембраны оказывается заряженной отрицательно за счет ионов хлора, аминокислот и других крупных органических анионов, которые через мембрану практически не проникают.
Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя, в этом участке возникает возбуждение, проявляющееся в быстром колебании мембранного потенциала и называемое потенциалом действия.
Потенциал действия можно зарегистрировать с помощью электродов, приложенных к внешней поверхности волокна (внеклеточное отведение), либо микроэлектрода, введенного в цитоплазму (внутриклеточное отведение) (рис. 22).
При введении микроэлектрода в нервное волокно сначала регистрируется потенциал величиной около 60 мВ. Это мембранный потенциал.
Потенциал действия представляет собой резкий скачок мембранного потенциала. При этом происходит не только изменение отрицательного потенциала покоя от величины - 60 мВ до нуля, но и переход за нулевую линию на несколько десятков милливольт, так что амплитуда потенциала действия превышает потенциал покоя и составляет приблизительно 90 мВ.
В потенциале действия различают его пик (спайк, по терминологии английских авторов) и следовые потенциалы. Во время пика мембранный потенциал меняет свой знак.
Причина возникновения потенциала действия - изменение ионной проницаемости мембраны. При раздражении проницаемость клеточной мембраны для ионов натрия повышается. стремится внутрь клетки, так как, во-первых, он заряжен положительно и его влекут внутрь электростатические силы, вовторых, концентрация его внутри клетки невелика. В покое клеточная мембрана была малопроницаемой для этого иона. Раздражение изменило проницаемость мембраны, и поток положительно заряженных ионов натрия из внешней среды клетки в цитоплазму значительно превысил поток ионов калия из клетки наружу. В результате произошло извращение потенциала мембраны (фаза деполяризации). Внутренняя поверхность мембраны стала заряженной положительно, а наружная вследствие потери положительно заряженных ионов натрия - отрицательно. В этот момент и регистрируется пик потенциала действия.
Повышение проницаемости мембраны для ионов натрия продолжается очень короткое время. Вслед за этим в клетке возникают восстановительные процессы, приводящие к тому, что проницаемость мембраны для ионов натрия вновь понижается, а для ионов калия возрастает. Поскольку ионы калия также заряжены положительно, выходя из клетки, они восстанавливают исходные отношения снаружи и внутри клетки (фаза реполяризации).
Накопления натрия внутри клетки при многократном возбуждении ее не происходит, потому что эвакуируется из нее постоянно за счет действия специального биохимического механизма, называемого «натриевым насосом». Есть данные и об активном транспорте калия с помощью «натрий-калиевого насоса».
Таким образом, согласно мембранной теории в происхождении биоэлектрических явлений решающее значение имеет избирательная проницаемость клеточной мембраны, обусловливающая разный ионный состав на поверхности и внутри клетки, а следовательно, и разный заряд этих поверхностей. Следует заметить, что многие положения мембранной теории все еще дискуссионны и нуждаются в углубленной разработке.
Итак, биоэлектрические свойства клеток, обусловленные неравномерным распределением минеральных ионов, играют ведущую роль в процессах клеточного возбуждения. Возникновение потенциала действия является наиболее характерным показателем возбуждения в клетках, тканях и органах. Поэтому электрофизиологические исследования широко применяются в экспериментальных лабораториях и в клиниках.
Значение регистрации биоэлектрических явлений. Регистрация электрических потенциалов в живом организме дает возможность судить об изменениях многих физиологических функций. Так как электрическая активность многих органов человека в норме имеет типичные и постоянные характеристики, методы электрофизиологии широко используются для диагностики болезней в практической медицине. Большие успехи достигнуты в тонком распознавании болезней сердца, нервной системы, мышц. Электрофизиологические методы сыграли важную роль в решении многих проблем космической физиологии. С помощью методов телеметрии оказалось возможным передавать информацию о состоянии сердечной мышцы, деятельности мозга, скелетной мускулатуры и других органов в условиях невесомости, перегрузок.
Изучение электрической активности центральной нервной системы легло в основу конструирования сложных счетно-решающих электронных машин.
Под электроэнцефалографией понимают запись биоэлектрических явлений, протекающих в головном мозге, преимущественно в коре больших полушарий головного мозга.
Для отведения биотоков от различных структур головного мозга используют различные конструкции электродов. В эксперименте на животных электроды можно ввести через кости черепа прямо в нужный участок головного мозга. Такие «вживленные» электроды долго удерживаются в мозгу специальным креплением и позволяют изучать электрическую активность определенных участков головного мозга при различных состояниях животного.
 Рис. 23.
Рис. 23.
В настоящее время возможна длительная регистрация электрических явлений даже в отдельных клетках мозга с помощью микроэлектродов.
При записи биотоков мозга человека - электроэнцефалограммы - пользуются обычно серебряными электродами, имеющими вид пластинки размером с двухкопеечную монету. Электроды на голове испытуемого человека укрепляются с помощью шлемов-сеток. Шлемы изготовляют из эластичных резиновых тяжей, натяжение которых регулируют. Шлемы, плотно прилегая к голове испытуемого, надежно удерживают электроды (рис. 23).
Запись биотоков мозга производится на приборах - электроэнцефалографах, имеющих разную конструкцию и включающих несколько усилителей биотоков, осциллографы и сложный пульт управления ими.
В настоящее время выпускают приборы, позволяющие регистрировать одновременно электрическую активность от 2 до 32 точек мозга и более.
Для точного анализа электрических явлений, возникающих в головном мозге и других органах, используют электронно-вычислительные машины.
Электрическая активность мозга человека носит ритмический характер. Электроды, расположенные на поверхности головы, дают возможность зарегистрировать биотоки сразу от многих клеток мозга, лежащих под ними. Поэтому общий характер энцефалограммы оказывается очень сложным. Вместе с тем удалось установить, что наиболее выраженных и часто встречающихся ритмов колебания электрической активности немного. Названия этих ритмов условно обозначены греческими буквами α, β, δ, ϑ (рис. 25, табл. 4).
Таблица 4
Ритмы электроэцефалограммы
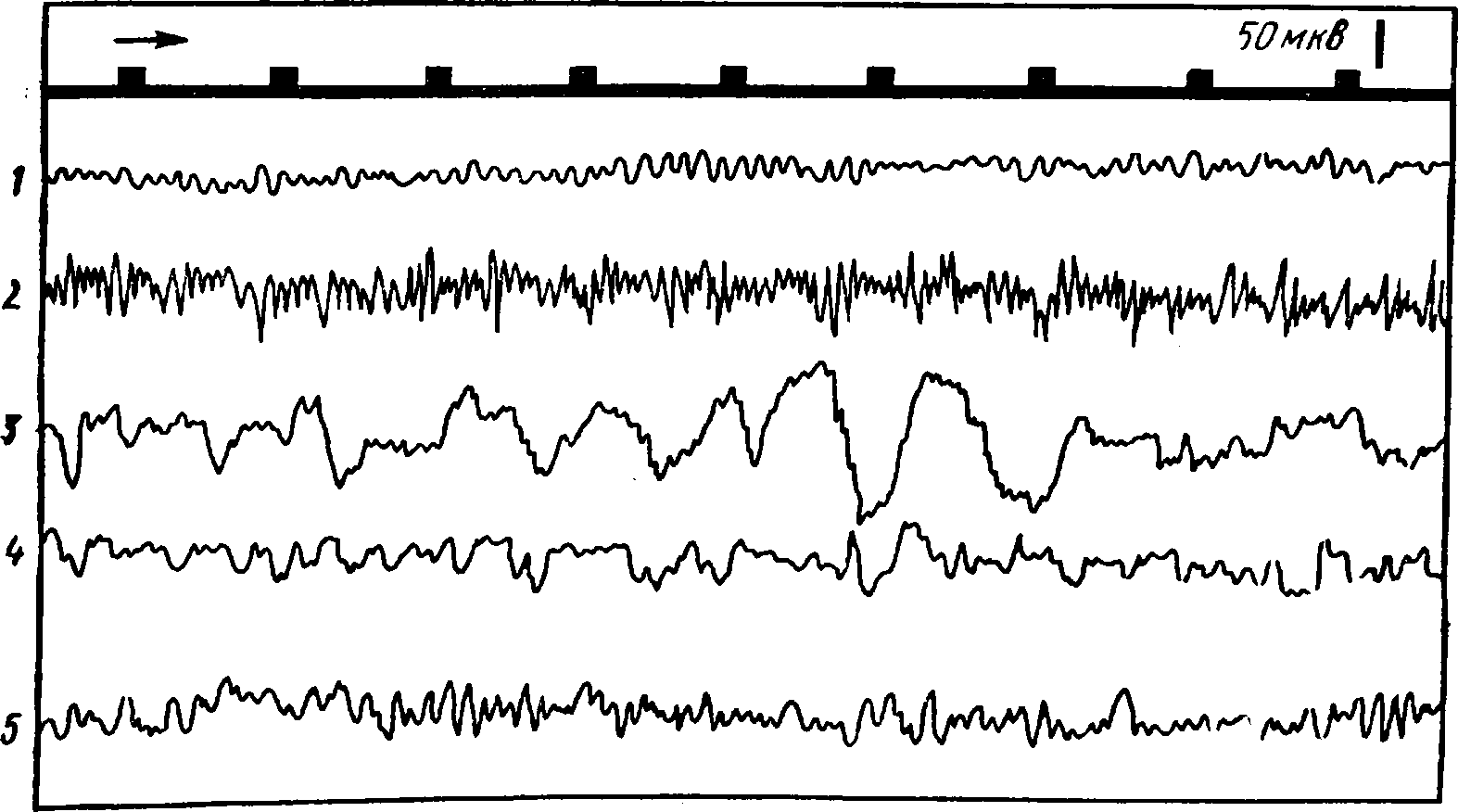 Рис. 25.
Рис. 25.
1 -альфа-ритм- 2 - бета-ритм; 3 - дельта-ритм; 4- тета-ритм; 5 -веретена; вверх у - отметка времени
При ограниченном поступлении центростремительных импульсов в исследуемый участок мозга обычно наблюдаются медленные волны с большим размахом колебаний. Если в кору поступают многочисленные импульсы, то клетки в этом участке могут находиться в разных стадиях возбуждения и общая электрическая активность над этим участком характеризуется частыми колебаниями с небольшой амплитудой типа бета-ритма. На рисунке 26 приведена электроэнцефалограмма с затылочной области коры (там располагается зрительная зона коры больших полушарий). На ней отчетливо виден переход альфа-ритма в бета-ритм и обратно при открывании и закрывании глаз, т. е. при увеличении и снижении потока центростремительных импульсов к зрительной зоне коры.
Современная техника позволяет регистрировать энцефалограмму у человека на расстоянии и даже в условиях космического полета.
Получил достаточно широкое распространение и метод исследования электрической активности мышц - электромиография.
 Для отведения биопотенциалов мышц человека используют накожные металлические электроды диаметром 10 мм. Электроды укрепляют на исследуемой мышце эластической манжетой; между ними и кожей находится обычно специальна
я паста, улучшающая контакт с телом и электропроводность. Колебания биопотенциалов мышц имеют также ритмический характер, только частота их и амплитуда значительно больше, чем при записи электроэнцефалограммы. Усиление мышечной активности сопровождается увеличением амплитуды и колебаний электромиограммы. При утомлении мышц частота колебаний, как правило, падает.
Для отведения биопотенциалов мышц человека используют накожные металлические электроды диаметром 10 мм. Электроды укрепляют на исследуемой мышце эластической манжетой; между ними и кожей находится обычно специальна
я паста, улучшающая контакт с телом и электропроводность. Колебания биопотенциалов мышц имеют также ритмический характер, только частота их и амплитуда значительно больше, чем при записи электроэнцефалограммы. Усиление мышечной активности сопровождается увеличением амплитуды и колебаний электромиограммы. При утомлении мышц частота колебаний, как правило, падает.
Рис. 26. Изменения электроэнцефалограммы затылочной области коры, показывающие переход от альфа-ритма к бета-ритму при открывании глаз (стрелка вверх) и восстановление альфа-ритма при закрывании глаз (стрелка вниз).
На рисунке 27 представлены электромиограммы трехглавой и двуглавой мышц плеча при работе напильником. На них отчетливо видно усиление колебаний при возбуждении мышц. Электромиография позволяет установить степень участия тех или иных мышц в выполняемом движении. Особенно важен этот метод для установления наличия паралича мышц при некоторых заболеваниях. Так, с помощью электромиографии можно обнаружить паралич дыхательных мышц при полиомиелите до того, как остановится . А это очень важно для принятия необходимых мер (перевод на управляемое с помощью специальной аппаратуры) с целью спасения жизни человеку.
 Рис. 27.
Электромиограммы трехглавой (1)
и двуглавой (2)
мышц
плеча у рабочего при опиловке.
Рис. 27.
Электромиограммы трехглавой (1)
и двуглавой (2)
мышц
плеча у рабочего при опиловке.
Данные электромиографии используются при создании активных протезов и управляемых манипуляторов.
«Искусственная рука» - одно из первых устройств, управляемых с помощью биотоков мышц. Это активный протез, воспроизводящий нормальные движения кисти. Такой активный протез имеет браслет для отведения биотоков мышц сохранившейся культи. На браслете укреплены металлические чашечки, заполненные токопроводящей пастой. Отводимые с помощью браслета мышечные токи усиливаются и поступают в специальный блок на протезе, где формируется управляющий сигнал. Сигнал приводит в действие миниатюрный двигатель, который обеспечивает движение протеза. Сам протез выполняется из пластических материалов, сходных по форме, размерам и цвету с нормальной рукой.
Передача возбуждения в синапсах
Возбуждение от одной нервной клетки к другой или от нервного волокна к мышечной или железистой клетке передается с помощью синап сов.
Аксоны большинства ней ронов, подходя к другим нервным клеткам, ветвятся и образуют многочисленные окончания на телах этих кле ток и их дендринах (рис. 28). Такие места контактов называют синапсами.
 Рис. 28.
Расположение синапсов на теле нейрона и на его дендритах (по Хаачу и Барру)
Рис. 28.
Расположение синапсов на теле нейрона и на его дендритах (по Хаачу и Барру)
Количество синапсов на теле одного нейрона достигает 100 и больше, а на дендритах одного нейрона - нескольких тысяч. Одно нервное волокно может образовать до 10 000 синапсов на многих нервных клетках.
Синапс имеет сложное строение (рис. 29). Он образован двумя мембранами - пресинаптической и постси наптической, между которыми есть синоптическая щель. Пресинаптическая мембрана находится на нервном окончании. Нервные окончания в центральной нервной системе имеют вид пуговок, колечек или бляшек. На теле пирамидной клетки насчитывается несколько тысяч нервных окончаний. Каждая синаптическая пуговка покрыта пресинаптической мембраной. Постсинаптическая мембрана находится на теле или дендритах нейрона, к которому передается нервный импульс. В пресинаптической области обычно наблюдаются большие скопления митохондрий.
Возбуждение через синапсы передается химическим путем с помощью особого — посредника, или медиатора, находящегося в синаптической бляшке. В синаптической бляшке медиатор находится в синаптических пузырьках, каждый из которых содержит до 2000 молекул медиатора. В разных синапсах вырабатываются разные медиаторы. Чаще всего в качестве медиатора служит ацетилхолин, адреналин или норадреналин в некоторых синапсах - глютаминовая кислота.
Каков же механизм передачи возбуждения в синапсах? Приход нервного импульса в пресинаптическое окончание сопровождается синхронным выбросом в синаптическую щель медиатора из синаптических пузырьков, расположенных в непосредственной близости от нее. Размеры этой щели очень малы (около 200 Å), и медиатор, быстро достигая постсинаптической мембраны, взаимодействует с ее веществом. В результате этого взаимодействия структура постсинаптической мембраны временно изменяется, проницаемость ее для ионов натрия повышается, что приводит к перемещению ионов и, как следствие, появлению возбуждающего постсинаптического потенциала. Когда этот потенциал достигает определенной величины, возникает распространяющее возбуждение - потенциал действия.
 Рис. 29.
Рис. 29.
1 - пресинаптическая мембрана; 2 - постсинаптическая мембрана; 3 - синаптичес кая щель; 4 - синаптические пузырьки; 5 - нейропротофибриллы; 6 - митохондрии.
Через очень короткое время (несколько миллисекунд) медиаторы разрушаются специальными ферментами.
В настоящее время подавляющее большинство нейрофизиологов признает существование в спинном мозге и в различных отделах головного мозга двух качественно различных типов синапсов - возбуждающих и тормозящих. Полагают, что в специализированных тормозящих нейронах, в нервных окончаниях аксонов вырабатывается особый медиатор, оказывающий тормозящее воздействие на последующий нейрон. Природа этого медиатора еще окончательно не установлена. В коре больших полушарий таким медиатором некоторые авторы считают гамма-амино-масляную кислоту.
Под влиянием приходящего по аксону тормозящего нейрона импульса в синаптическую щель выделяется медиатор, который вызывает специфические изменения в постсинаптической мембране. Суть этих изменений состоит в том, что медиатор торможения, взаимодействуя с веществом постсинаптической мембраны, увеличивает ее проницаемость для ионов калия и хлора. Внутри клетки относительно увеличивается число анионов. В результате происходит не снижение величины внутреннего заряда мембраны (как это имеет место в возбуждающем синапсе), а повышение внутреннего заряда постсинаптической мембраны. Это ведет к возникновению Главная Анатомия человека
Токи покоя Наличие разности потенциалов можно обнаружить, если к поврежденной мышце приложить электроды осциллографа таким образом, чтобы один из них... История изучения человека Анатомия физиология человека Место человека в природе Клетка Обмен веществ клетки Деление... Обмен веществ Регуляция деятельности организма Нервная система Рефлекс и рефлекторная дуга Безусловные и условные рефлексы Кровь Плазма крови Реакция крови...

1. Понятия и общие свойства возбудимых тканей Возбудимые ткани: Возбудимые ткани: мышечная, нервная, железистая. Все клетки и ткани живых систем в процессе эволюции приобрели способность отвечать на действие раздражителя специфической и неспецифической реакцией. Раздражимость Раздражимость – это способность органов, тканей, клеток активно отвечать на воздействие какой-либо формой деятельности – обменом, ускорением деления, выбросом секрета, движением, электрическим импульсом. Возбудимость Возбудимость – процесс временной деполяризации мембраны клеток со специфической ответной реакцией при котором мышечная ткань сокращается, железистая – выделяет секрет, нервная – проводит нервный импульс.


Классификация раздражителей. 1. По энергетической природе:физические,химические,биологические. эндогенные (внутренние) 2. По месту нахождения раздражителя: эндогенные (внутренние); экзогенные (внешние) экзогенные (внешние). 3. По силе раздражителя: подпороговой силы; пороговой силы пороговой силы – раздражитель минимальной величины, вызывает ответную реакцию у возбудимой ткани; сверхпороговой сверхпороговой. 4. По биологическому значению:адекватные неадекватные

Условия возникновения возбуждения. Возбуждение Возбуждение – это сложный физиологический процесс перехода клетки или ткани от состояния покоя к выполнению специализированных функций. –Наличие реактивной ткани. –Присутствие раздражителя пороговой силы. –Градиент раздражения (закон градиента) – скорость нарастания силы раздражителя, она должна быть очень высокой (мгновенной). –Аккомодация – это приспособление возбудимой ткани к медленному нарастанию силы раздражителя. При этом возбуждение может и не наступить. –Время действия раздражителя. Чем больше сила раздражителя (не превышая порог), тем меньше времени он должен действовать, чтобы вызвать возбуждение.

Показатели измерения возбудимости –Реобаза –Реобаза – пороговая сила раздражителя – электрического тока –Полезное время –Полезное время – это минимальное время действия раздражителя пороговой силы. –Хронаксия –Хронаксия – это минимальное время действия раздражителя удвоенной реобазы (мс), способного вызвать ответную реакцию. Чем выше возбудимость ткани, тем меньше хронаксия.

Мембранный потенциал (потенциал покоя) Мембранный потенциал (потенциал покоя) – это разница зарядов между наружной и внутренней сторонами плазматической мембраны. двумя факторами: Согласно мембранно-ионной теории, мембранный потенциал обусловлен двумя факторами: 1. Ассиметричным расположением ионов 1. Ассиметричным расположением ионов между внутренним содержимым клетки и внеклеточной жидкостью. 2. Избирательной проницаемостью мембраны клетки 2. Избирательной проницаемостью мембраны клетки – это обеспечивают специальные отверстия в ней – поры или «каналы» (до 500 шт. на мм 2 поверхности). Na + Главным катионом межклеточной жидкости является Na +,которого в 15 раз больше, чем в цитоплазме клетки – это создаёт положительный заряд наружной части мембраны. 2. Биоэлектрические потенциалы в возбудимых тканях

Мембранный потенциал Главными катионами внутриклеточной жидкости (цитоплазмы) ионы К + Главными катионами внутриклеточной жидкости (цитоплазмы) являются ионы К +, которого в 40 раз больше, чем в межтканевой жидкости. Согласно законам осмоса и диффузии, ионы К + по градиенту концентрации выходят через поры мембраны в межтканевую жидкость, увеличивая тем самым положительный заряд наружной стороны мембраны. Отрицательный заряд внутренней стороны мембраны создаётся за счёт анионов: НСО 3 -, NaHCO 3 -, органических кислот: уксусная, пропионовая, аминокислоты.



Потенциал действия Na + в 500 раз В результате действия раздражителя открываются натриевые каналы и увеличивается пропускная способность мембраны клетки для ионов Na + в 500 раз. Наблюдается лавинообразный поток движения катионов Na + внутрь клетки. Всё это приводит к деполяризации мембраны с последующей фазой смены полярности. Внешняя сторона мембраны становится электроотрицательной. В результате этого возникают биотоки и совершается действие.





Фазовые изменения возбудимости или развитие процесса возбуждения развитие процесса возбуждения –Латентный период (скрытый) –Латентный период (скрытый) – предшествующий видимому проявлению возбуждения ткани (сокращение, проведение нервного импульса, выделение секрета). Продолжительность – 0,01 с. –Фаза абсолютной рефрактерности- –Фаза абсолютной рефрактерности- полная невосприимчивость к повторным раздражениям – совпадает с восходящей частью пика потенциала действия. –Фаза относительной рефрактерности (пониженная возбудимость) –Фаза относительной рефрактерности (пониженная возбудимость) – возбудимость ткани постепенно восстанавливается до исходного уровня. –Фаза экзальтации (гиперполяризации) –Фаза экзальтации (гиперполяризации) – повышенной возбудимости.

Фазовые изменения возбудимости на примере одиночного сокращения мышечной ткани. 1. Латентный период. 2. Фаза укорочения 2. Фаза укорочения (абсолютной рефрактерности). 3. Фаза расслабления 3. Фаза расслабления (относительной рефрактерности) 4. Фаза экзальтации 4. Фаза экзальтации (гиперполяризации) – повышенной возбудимости

3. Строение, свойства и функции мышечной ткани. различают три вида мышечной ткани: У позвоночных животных различают три вида мышечной ткани: 1. скелетная поперечнополосатая мышечная ткань 1. скелетная поперечнополосатая мышечная ткань (произвольная). 2. поперечнополосатая сердечная ткань 2. поперечнополосатая сердечная ткань (непроизвольная). 3. гладкая мышечная ткань 3. гладкая мышечная ткань внутренних органов, кровеносных сосудов, кожи (непроизвольная). Скелетные мышцы сарколеммойсаркоплазма (протоплазма- ээтическое вещество) миофибриллы митохондрий Скелетные мышцы состоят из мышечных волокон (клетки) диаметром от 20 до 100 мкм длинной см. Каждое волокно покрыто оболочкой – сарколеммой, внутри – саркоплазма (протоплазма- ээтическое вещество) и многочисленные тонкие нити – миофибриллы, количество которых достигает шт. диаметром 0,5-2 мкм. Между миофибриллами большое количество митохондрий для снабжения мышц энергией.


Миофибриллы Миофибриллы имеют исчерченность – это чередование тёмных и светлых сегментов. протофибрилл анизотропные изотропные В структуру миофибрилл входят протофибрилл (филаменты) в которых идет чередование молекул белка миозина –анизотропные, темные диски и молекул белка актина – изотропные, светлые диски. саркомер. теории «скольжение нитей» Х. Хаксли и А. Хаксли тропомиозина, тропинина и ионов кальция. Функциональной и структурной единицей является саркомер. Это повторяющиеся в миофибриллах блоки светлых и тёмных дисков отделённых друг от друга £-пластинками. Механизм сокращения мышц согласно теории «скольжение нитей» Х. Хаксли и А. Хаксли – есть перемещение актиновых нитей вдоль миозиновых к центру саркомера, при активном участии белков тропомиозина, тропинина и ионов кальция.


Схема строения поперечнополосатой мышечной ткани: 1 - эндомизий; 2 - мышечные волокна; 3 - сарколемма; 4 - пучки миофибрилл; 5 - миофибрилла; 6 - анизотропный диск; 7 - изотропный диск; 8 - ядра; 9 - кровеносные капилляры; 10 - соединительнотканные клетки эндомизия; 11 - моторное нервное волокно; 12 - моторное нервное окончание.


А. Актиновая и миозиновая нити на продольном сечении волокна. Б. Они же на его поперечном сечении. Когда Са 2+ связывается с тропонином, тропомиозин попадает в желобок между двумя мономерами актина, обнажая участки прикрепления поперечных мостиков Механизм мышечного сокращения. Действие Ca 2+ во время активации миофибриллы.



Свойства мышечной ткани 1. Возбудимость 1. Возбудимость - свойство мышечной ткани отвечать на действие раздражителя специфическим изменением проницаемости мембраны, возникновением разности потенциалов и электродвижущей силы (ЭДС). 2. Проводимость 2. Проводимость – возбуждение распространяется по всему мышечному волокну и не переходит на рядом лежащие, т. к. сарколемма служит изолятором.

3. Сократимость 3. Сократимость – основная функция мышечной ткани. При этом она укорачивается, утолщается, изменяя свои линейные размеры. изотоническое сокращение изотоническое сокращение – без изменения тонуса; изометрическое сокращение изометрическое сокращение – без изменения линейных размеров мышцы; ауксотоническое сокращениеауксотоническое сокращение – смешенное сокращение при котором изменяется и длинна и тонус мышц. 4. Эластичность 4. Эластичность – когда после прекращения действия деформирующей силы – мышечная ткань принимает первоначальные размеры.

Виды сокращения мышц 1. Одиночное сокращение 1. Одиночное сокращение – возникает как ответ на одиночное кратковременное действие раздражителя. 2. Тоническое сокращение 2. Тоническое сокращение - это сильное длительное сокращение мышцы при действии раздражителя высокой частоты: зубчатый тетанус зубчатый тетанус – возникает при частоте подачи раздражителя 5-15 Гц в секунду. гладкий тетанус гладкий тетанус – возникает при частоте подачи раздражителя более 20 Гц в секунду.


Работа, сила и утомление мышц Работа Работа – это произведение величины поднятого груза на высоту его поднятия. A=P*h Динамическая работа Динамическая работа - при которой происходит перемещение груза и движение костей в суставах. Статическая работа Статическая работа – происходит при изометрическом сокращении мышц. В этом случае внешняя работа не совершается.

Сила мышц физиологическая площадь поперечного сечения Сила мышц – величина максимально поднятого груза и зависит от количества и толщины мышечных волокон, т. е. решающее значение имеет физиологическая площадь поперечного сечения мышечных волокон. Физиологическое поперечное сечение (поперечник) совпадает с анатомическим только в мышцах с продольно расположенными волокнами. У мышц с косым направлением мышечных волокон физиологический поперечник больше анатомического. Поэтому и сила мышц с косыми волокнами всегда больше. абсолютная сила мышц Сравнительным показателем силы различных мышц является абсолютная сила мышц – это величина максимально поднятого груза делённая на квадрат поперечного сечения мышцы.


Слева: мышца веретенообразной формы, её анатомический и физиологический поперечники совпадают. То же характерно для мышц лентовидной формы. Справа: мышца двоякоперистой формы, её физиологический поперечник значительно больше анатомического (анатомический поперечник не показан).

Утомление мышц Утомление мышц – это временное снижение или полная потеря работоспособности после длительных нагрузок. Причиной утомляемости является: истощение запасов медиатораистощение запасов медиатора в синапсах и АТФ, креотинфосфата (КФ), гликогена в мышцах (энергетический материал); отравление мышц продуктами метаболизмаотравление мышц продуктами метаболизма – накопление в мышцах молочной, угольной и фосфорной кислот и др., что вызывает обратимые изменения сократительных белков мышечной ткани

Энергия (химизм) мышечного Сокращения две фазы: Энергия, необходимая для работы мышц образуется в результате сложных химических процессов, протекающих в две фазы:анаэробная АТФ=АДФ+Н 3 РО 4 +Q Р+(С 6 Н 12 О 6)n Гексозофосфат (ГФ) гликоген ГФМолочная кислота+Н 3 РО 4 +Q Креатин+Н 3 РО 4 Креатинфосфат (КФ) аэробная фаза аэробная фаза – идёт окисление молочной кислоты до СО 2 и Н 2 О около 20%, а 80% идёт на ресинтез гликогена. Креатинфосфат (КФ)Креатин+Н 3 РО 4 +Q АДФ+Н 3 РО 4 =АТФ

Свойства гладкой мышечной ткани Гладкая мышечная ткань находится во внутренних органах, в кровеносных сосудах и коже. миоциты Структурными и функциональными элементами являются одноядерные мышечные клетки миоциты веретенообразной формы, соединенные между собой дисками (нексусы). Миофибриллы мышечной клетки Миофибриллы мышечной клетки размещаются параллельно друг другу. Актиновые и миозиновые нити распределены неравномерно – поэтому нет исчерченности.

Свойства гладкой мышечной ткани 1. Возбудимость и проводимость 1. Возбудимость и проводимость ниже, чем у скелетных мышц; 2.2. Удлинен латентный (скрытый) период 2.2. Удлинен латентный (скрытый) период до 1 сек.; 3. Сократимость миофибрил 3. Сократимость миофибрил осуществляется с участием белков тропомиозина, тропинина и ионов Са ++. Продолжительность до 100 сек.; 4. Пластический тонус 4. Пластический тонус – гладкие мышцы способны изменять линейные размеры (растягиваться) не изменяя своего тонуса. 5. Автоматизм 5. Автоматизм – способность гладкой мышечной ткани сокращаться под воздействием импульсов, которые зарождаются в ней самой (собственная интрамуральная нервная система – нервные ганглии, которые самостоятельно генерируют потенциал действия).

3. Физиология нервной ткани. Эволюция нервной системы тесно связанна с развитием двигательной функции живых организмов и развивалась путём усовершенствования способов передачи возбуждения, дифференцировки и концентрации клеточных структур. различают три типа нервной системы: По этим признакам различают три типа нервной системы: диффузную, ганглиозную и трубчатую. 1. Диффузная нервная система 1. Диффузная нервная система имеется у простейших многоклеточных животных (гидра пресноводная). Она состоит из малодифференцированных нервных клеток, отвечающих на действие раздражителя общей однотипной реакцией.

2. Ганглиозная нервная система. 2. Ганглиозная нервная система. Нейроны концентрируются в определённых местах тела животного, образуя узлы-ганглии. Одновременно происходит специализация нервных клеток: сенсорные, ассоциативные и эффекторные. Эта нервная система появляется у червей и достигает совершенства у представителей типа членистоногих. 3. Трубчатая нервная система. 3. Трубчатая нервная система. У позвоночных животных нейроны образуют трубку, расширенную в передней части в виде пузыря из которого формируется головной мозг.

Нервная система Нервная система осуществляет связь с внешней средой, регулирует работу всех органов и систем живого организма, адаптируя их функцию к изменяющимся условиям внешней среды. Материалом для построения нервной системы служит Материалом для построения нервной системы служит нервная ткань, которая состоит из двух компонентов: нейронов и клеток нейроглии. Основными функциональными элементами ЦНС Основными функциональными элементами ЦНС являются нейроны, их примерно 50 млрд и это составляет 10-15% от общего количества клеточных элементов нервной системы.

Нервная клетка состоит дендритов (лат. dendron - дерево) аксона (нейрит) аксональные терминали Нервная клетка состоит из тела сомы, (перикарион) и отростков разного типа: дендритов (лат. dendron - дерево) - многочисленные короткие отростки (чувствительные) через катрые сигналы поступают в нервную клетку; аксона (нейрит)– длинный отросток, всегда один, выполняет эфферентную функцию. Окончанием у этих отростков служат аксональные терминали. Функции нейрона: восприятие сигналов, хранение и переработка информации, передачи нервных импульсов к эффекторам.


Классификация нейронов 1. По форме: 1. По форме: пирамидные, круглые, звёздчатые, овальные. Размеры от 5 до 150 мкм. 2. По количеству отростков: униполярныеуниполярные - одноотростковые нейроны, псевдоуниполярныепсевдоуниполярные - сенсорные нейроны спинного мозга, биполярныебиполярные - двухотростковые нейроны, мультиполярныемультиполярные -многоотростковые нейроны. 3. По выполняемой функции: сенсорные нейронысенсорные нейроны - чувствительные (афферентные), ассоциативныеассоциативные – вставочные, промежуточные более мелкие различной формы. эффекторныеэффекторные – двигательные, секреторные и т. д.

Клетки нейроглии функции Клетки нейроглии составляют основную массу нервной ткани (85-90%) – это эпиндимоциты, астроциты (протоплазматические, волокнистые), олигодендроциты, микроглия. Они заполняют всё пространство между нейронами и выполняют следующие функции: опорно-структурная, обмен веществ, трофическая, буферная (постоянная концентрация ионов К +), защитная (фагоцитоз).



Нервные волокна Нервные волокна – это отростки нейронов различной длинны, часто соединяющихся между собой, образуя нервные стволы. Мякотные (миелиновые) нервные волокна Мякотные (миелиновые) нервные волокна выполняют чувствительную и двигательную функции. Иннервируют в основном скелетную мускулатуру. Скорость проведения возбуждения м/сек. Безмякотные (безмиелиновые) нервные волокна Безмякотные (безмиелиновые) нервные волокна входят в состав вегетативной нервной системы (ВНС) и иннервируют внутренние органы. Скорость проведения возбуждения 0,5-3,0 м/сек.

Функциональная значимость структурных элементов нервного волокна: мембрана осевого цилиндрамембрана осевого цилиндра, толщина А 0, обеспечивает возникновение возбуждения и передачу его по нерву; миелиновая оболочкамиелиновая оболочка имеет липидную природу(фосфолипид-диэлектрик). Является изолятором и выполняет трофическую функцию, обеспечивает высокую скорость передачи нервного импульса и формирует канал роста аксона при его повреждении (регенерация); перехват Ранвьеперехват Ранвье – место, где возникает возбуждение; швановская клеткашвановская клетка – продуцирует миелин; нейрофибриллынейрофибриллы обеспечивают рост нервного волокна.

Строение миелинового нервного волокна 1 осевой цилиндр 1 осевой цилиндр (отросток нервной клетки). В миелиновом волокне он всего один, располагается в центре и значительно больше по диаметру, чем в безмиелиновом волокне. 2 миелиновый слой оболочки волокна. 2 миелиновый слой оболочки волокна. Это несколько слоев мембраны шваннов-ских клеток (леммоцитов), концентрически закрученных вокруг осевого цилиндра. Фактически это сильно удлиненный мезаксон. 3 цитоплазма леммоцита 3 цитоплазма леммоцита. 4 ядро леммоцита 4 ядро леммоцита: вместе с цитоплазмой оттеснено к периферии волокна и образует нейролемму наружный слой оболочки миелинового волокна. 5 базальная мембрана 5 базальная мембрана, окружающая волокно. Схема по Т.Н. Радостиной, Ю.И.Афанасьеву, Т. С. Румянцевой

Схема строения миелинового нервного волокна 1 – 1 – осевой цилиндр, 2 – 2 – миелиновый слой оболочки волокна, 3 – 3 – перехват Ранвье, 4 – 4 – клетка Швана (леммоцита), 5 – 5 – метахондрии, 6 – 6 – нейроплазма, 7 – 7 – ядро клетки Швана, 8 – 8 – нейрофибрилы.

Закономерности проведения возбуждения по нервному волокну 1. Проведение импульсов по нервному волокну 1. Проведение импульсов по нервному волокну возможно только в условиях анатомической и физиологической непрерывности. 2. Возбуждение по нервному волокну 2. Возбуждение по нервному волокну распространяется в обе стороны с одинаковой силой и скоростью. 3. По нервному волокну импульсы изолированно 3. По нервному волокну импульсы распространяются изолированно, не переходят с одного нерва на другой. 4. Нервное волокно в качестве энергии 4. Нервное волокно в качестве энергии использует АТФ и КФ. Однако интенсивность обмена веществ ничтожно мала. Это подтверждает небольшое количество образованного тепла. 5. Относительная не утомляемость нервного волокна 5. Относительная не утомляемость нервного волокна объясняется тем, что при возбуждении тратится небольшое количество энергии.

6. Проведение потенциала действия по нервному волокну 6. Проведение потенциала действия по нервному волокну осуществляется последовательно с определённой скоростью без затухания. В основе объяснения этого механизма лежит теория местных токов А. Ходжкина. Согласно этой теории, в результате раздражения изменяется ионная проницаемость мембраны нервного волокна и появляются разнозаряженные участки (+-) между которыми возникают местные круговые токи. Эти токи движутся вдоль нервного волокна от участка к участку.

Синапс Синапс (греч. sinapsis – соединение, связь) – место контакта нервного волокна с эффектором. Классификация синапсов. 1. В зависимости от локализации 1. В зависимости от локализации – центральные и периферические синапсы; 2. По способу передачи возбуждения 2. По способу передачи возбуждения – химические, электрические, смешанные синапсы; 3. По типу контакта: 3. По типу контакта: межнейронные (аксодендритический, аксосоматический и аксо-аксональный синапсы), нервно- мышечные, рецепторно-нейронные и нервно-железистые синапсы. 4. Возбуждающие синапсы 4. Возбуждающие синапсы - для передачи нервного импульса в них используются медиаторы: ацетилхолин, норадреналин, дофамин, аспарагиновая, глютаминовая кислоты, серотонин; 5. Тормозные синапсы 5. Тормозные синапсы – где генерируется тормозной постсинаптический потенциал (ТПСП) и используются медиаторы: гамма-аминомасляная кислота и аминокислота глицин.
Механизм синаптической передачи возбуждения Под действием нервного импульса из синаптических пузырьков конца аксона освобождается медиатор и впрыскивается (экзоцитоз) в синаптическую щель. В результате этого происходит перемещение ионов (поток Nа + внутрь превышает поток К + наружу, в клетку поступают ионы Са ++) и возникает возбуждающий постсинаптический потенциал (ВПСП).

В основе межнейронных связей лежит взаимодействие процессов возбуждения и торможения. Постсинапээтическое торможение возникает вследствие снижения возбудимости сомы и дендритов нейрона. В основе этого торможения лежит возникновение гиперполяризующего тормозного постсинаптического потенциала (ТПСП) в синаптических бляшках. Пресинапээтическое торможение возникает при уменьшении или прекращении высвобождения медиатора из пресинаптических нервных окончаний.

ПРЕДМЕТ ФИЗИОЛОГИЯ Представление о физиологических и функциональных системах организма
ФИЗИОЛОГИЯ – это биологическая дисциплина,
изучающая функции целостногоорганизма, отдельных
физиологических систем, тканей,
клеток, а также их взаимодействие и
регуляцию.Большой раздел физиологии отведен
изучению физиологических и
функциональных систем.
Физиологическая система – это постоянная
совокупность различных органов,
объединенных какой-либо общей функцией.
Функциональная система – это временная
совокупность органов,которые относятся к разным анатомическим
и физиологическим структурам, но
обеспечивают выполнение особых форм
физиологической деятельности и
определенных функций. Функциональная
система обладает свойствами:
1. саморегуляция;
2. динамичность (распадается после
достижения желаемого результата);
3. наличие обратной связи.Благодаря этому организм работает как единое целое. Особое
место в физиологии уделяется гомеостазу.
Гомеостаз – это совокупность биологических реакций,
обеспечивающих постоянство внутренней среды организма
(кровь, лимфа, межклеточная и цереброспинальная жидкость).ЛЕКЦИЯ №1. Физиология возбудимых
тканей.
1. Физиологическая характеристика
возбудимых тканей. Параметры возбудимости.
К возбудимым тканям относят: нервную,
мышечную, железистую. Возбудимость - это
способность ткани под действием раздражителей
переходить из состояния функционального покоя
в состояние возбуждения.Раздражители – это факторы внешней и внутренней
среды, действующие на возбудимую ткань.
Классификация раздражителей.
а) по природе раздражители бывают: физические,
химические, физико-химические, биологические,
социальные.
б) по физиологической значимости: адекватные и
неадекватные. Адекватными называют те раздражители, к
восприятию которых данная ткань приспособлена в
процессе своего эволюционного развития.(например, для мышечной ткани адекватными
являются нервные импульсы; для рецепторов
глаза – световой раздражитель, для
рецепторов уха – звуковые колебания и т.д.).
Неадекватные – это неестественные
раздражители, к восприятию которых ткань
не приспособлена (к примеру, действие
электрического тока на мышцу или
механический удар по глазу).в) по биологической значимости
раздражители делят на положительные
(оказывающие благотворный эффект на
жизнедеятельность ткани) и отрицательные
(разрушающие жизнеспособность тканей).
г) по силе раздражители делят на:
подпороговые, пороговые, сверхпороговые.Подпороговые раздражители – это те
раздражители, сила которых меньше пороговой
(они не вызывают генерацию потенциала
действия). Пороговой силой называют
наименьшую силу раздражителя, которая
вызывает возбуждение ткани (генерацию в ней
потенциала действия). Сверхпороговые - это
раздражители, величина которых больше
пороговой.Общие свойства возбудимых тканей:
1. Возбудимость – способность живой ткани отвечать на
действие раздражителя изменением физиологических
свойств и возникновением процесса возбуждения.
Существуют силовые и временные показатели
возбудимости
К силовым показателям возбудимости относят
пороговую силу раздражителя (т.е., наименьшую силу
раздражителя, вызывающую возбуждение). Для
электрического тока введен специальный термин –
реобаза.Реобаза – это наименьшая сила электрического тока,
которая вызывает возбуждение ткани.
К временным показателям возбудимости относят:
полезное время раздражения и хронаксию. Полезное
время – это наименьшее время, в течение которого
электрический ток силой в одну реобазу вызывает
возбуждение ткани. Хронаксия – это наименьшее
время, в течение которого электрический ток силой в
две реобазы вызывает возбуждение ткани.
Зависимость силы от времени действия
Р – реобаза – это минимальная силатока, вызывающая возбуждение
ПВ – полезное время – минимальное
время
действия
раздражающего импульса силой в
одну реобазу, необходимое для
возбуждения.
Хр – хронаксия - минимальное
время действия раздражающего
импульса силой в 2 реобазы
необходимое для возникновения
ПД.В медицине, в частности в неврологический
практике, с диагностической целью определяется
хронакия мышц и двигательных нервов. Для этого
используется
специальный
прибор
–
хронаксиметр,
состоящий
из
источника
постоянного тока, набора сопротивлений и
приспособления для дозировки времени действия
тока.
Повреждение периферических нервов ведет к
резкому удлинению хронаксии
Методика хронаксиметрии помогает поставить
ранний диагноз или выявить эффективность
лечения.2. Лабильность или функциональная
подвижность – это способность
возбудимой ткани реагировать на
раздражение с определенной скоростью.
Мерой лабильности является
максимальное число импульсов,
возникающих в тканях в единицу
времени (в 1 сек.).3. Проводимость – способность ткани передавать
возбуждение от места раздражения по длине
возбудимой ткани.
4. Рефрактерность – временное снижение
возбудимости ткани в процессе его возбуждения
(бывает абсолютной, когда нет ответа ни на какой
раздражитель и относительной – при этом
возбудимость восстанавливается и ткань способна
реагировать на сильные (сверхпороговые)
раздражители.ЛЕКЦИЯ №2. Биоэлектрические явления
в возбудимых тканях. Природа потенциала
покоя (ПП) и потенциала действия (ПД).
О состоянии покоя в возбудимой ткани говорят в
том случае, когда на ткань не действует
раздражитель из внешней или внутренней среды.
При этом отмечается постоянный уровень
метаболизма, нет видимых проявлений функции
ткани.Основные формы активного состояния
возбудимой ткани – это возбуждение и
торможение.
Возбуждение – это активный физиологический
процесс, который возникает в ткани под
действием раздражителя.
При этом изменяются её физиологические
свойства, наблюдаются функциональные
отправления (в нерве возникает нервный
импульс, мышца сокращается).Торможение – это также активный
процесс, который возникает при
действии раздражителей на ткань.
Торможение проявляется в
подавлении (ослаблении)
возбуждения.История открытия электрических
явлений в возбудимых тканях
В конце XVIII века профессор
Болонского университета
Луиджи Гальвани провел ряд опытов,
благодаря которым появилось
представление о существовании
«животного электричества».В историю медицины вошли два
классических опыта Л.Гальвани. В первом
опыте ученый случайно использовал два
металла: препарат задних лапок лягушки он
подвешивал на медном крючке к железным
перилам балкона. Он заметил, что при
соприкосновении
мышц
препарата
с
железными перилами балкона
они
сокращаются.Ученый предположил, что причиной
сокращения мышц является электричество,
возникающее в живых тканях («животное»
электричество). Другой итальянский
ученый, физик Вольта, повторив опыт
Гальвани, пришел к заключению, что
причиной сокращения мышц являются
токи, возникающие между двумя
разнородными металлами: медью и
железом. И оказался прав.Однако Л.Гальвани доказал существование «животного»
электричества, выполнив свой второй опыт (без участия
металлов). Для этого он использовал нервно-мышечный
препарат. Повредив икроножную мышцу в проксимальной
части, он, с помощью стеклянного крючка, набрасывал
седалищный нерв на мышцу таким образом, чтобы он
одновременно прикоснулся к поврежденному и
неповрежденному участкам мышцы. При этом
икроножная мышца сокращалась. Причиной
возникновения нервного импульса явилась разность
зарядов: поврежденный участок мышцы
электроотрицателен (-) по отношению к неповрежденному
участку (+).В настоящее время появились методики
исследования,
которые
позволяют
регистрировать биопотенциалы даже на уровне
отдельных клеток (микроэлектродная техника).
Но еще задолго до появления этих методов
стало понятно, что «животное электричество»
обусловлено процессами, происходящими на
клеточной мембране.
Регистрация мембранного потенциала
МкЭ –микроэлектр
од
РЭ –
референтный
электрод
Клеточная мембрана
- это тонкая (6-10 нм) пластинка, котораясостоит из липидов и белков. Содержание
липидов – около 40%, белков – 60%;
- на внешней поверхности мембраны
имеется небольшое количество (5-10%)
углеводов (гликопротеидов, гликолипидов),
которые выполняют рецепторную и
защитную функцию.ФУНКЦИИ КЛЕТОЧНЫХ МЕМБРАН
Барьерная.
Рецепторная.
Транспортная.
Выработка биологически активных веществ.
Создание электрического заряда клетки.
Природа ПП и ПД
Потенциал покоя (ПП) – это относительностабильная разность электрических зарядов
между наружной и внутренней
поверхностью мембраны.
Её величина составляет 30-90 мВ.
Согласно
мембранно-ионной
теории
(Бернштейн, Ходжкин, Хаксли, Катц)
причиной разности зарядов на мембране
является
неодинаковая
концентрация
анионов и катионов внутри и вне клетки.Концентрация К+ внутри клетки в 30-40 раз больше,
чем вне клетки.
Концентрация Na+ вне клетки в 10-12 раз больше, чем
в клетке.
Ионов Сl- вне клетки в 30-50 раз больше, чем внутри
клетки.
В клетке содержатся также крупномолекулярные анионы
(белки – глутамат, аспартат, органические фосфаты).
ФАКТОРЫ, ФОРМИРУЮЩИЕ МП
ИОННАЯ АСИМЕТРИЯКонцентрационный градиент калия
= 20-40 p
1.
Концентрационный градиент натрия
= 8-10p
Ионы распределены неравномерно по обе стороны
мембраны за счет:
1. Неодинаковой проницаемости для них клеточной
мембраны;
2. Работы ионных насосов, которые транспортируют
ионы в клетку и из клетки против концентрационного и
электрического градиентов с затратой энергии АТФ.
2.Полупроницаемость мембраны
Na++
+
+
- -
+ + + +
- - -
Белок- K+
+
-
+
+Формирование потенциала покоя (ПП). Мембрана клетки
в состоянии покоя заряжена отрицательно изнутри и положительно
снаружи.
Проницаемость мембраны в покое для К+ в 25 раз выше, чем для
Na+.
По закону диффузии К+ выходит из клетки (т.к. его концентрация
внутри клетки в 40 раз больше, чем вне клетки). Органические
анионы из-за своих больших размеров выйти из клетки не могут и
они создают отрицательный заряд на внутренней поверхности
мембраны. Таким образом, главным ионом, который создает ПП
является ион К+.Формирование потенциала действия (ПД).
Потенциал действия выражается в быстром
колебании мембранного ПП при действии
раздражителя.
ПД обеспечивает передачу сигналов между
нервными клетками, в ЦНС, рабочих органах,
мышцах. Величина ПД составляет 80-130 мВ,
длительность – 0,5-1 мс.ПД включает:
– Фазу деполяризации (т.е. уменьшение мембранного потенциала до нуля);
1.
- Инверсии (изменение знака заряда на обратный: внутренняя поверхность
мембраны приобретает положительный заряд, наружная – отрицательный);
3.- Фазу реполяризации – восстановление первоначального заряда мембраны
(минус изнутри, плюс – снаружи);
4.- Следовые потенциалы (следовая деполяризация и следовая
гиперполяризация)
2.мВ
+30
3
4
0
2
Екр
1
Е0
-80
5
6
7Механизм формирования ПД:
При возбуждении клетки происходит активация Na-ых каналов. По
концентрационному градиенту ионы Na устремляются внутрь
клетки, где его концентрация меньше. При этом отрицательный
заряд внутренней поверхности мембраны уменьшается
(деполяризация). Затем разность зарядов возникает с обратным
знаком, так как число катионов в клетке превосходит число
анионов (инверсия, перезарядка).Через доли секунды (0,5-2 мсек) рост ПД прекращается,
вследствие инактивации Na-ых каналов и прекращения
поступления Na в клетку. При этом открываются К+-вые каналы.
Ионы К+ выходят из клетки по концентрационному и
электрическому градиентам. Вследствие этого восстанавливается
мембранный ПП (реполяризация).
Таким образом, главную роль в возникновении ПД играют ионы
Na+.Наряду с пассивным транспортом ионов
(диффузия), существует активный механизм
поддержания разности концентраций ионов
внутри и вне клетки.
Им является «натрий-калиевый насос».
Белки-переносчики (Na-K-АТФаза), расщепляя
молекулу АТФ, используют ее энергию на
перенос
ионов
(Na
и
К)
против
концентрационного
и
электрического
градиентов.
НАТРИЙ – КАЛИЕВЫЙ НАСОС
активный транспорт ионов натрия и калияпротив концентрационного градиента с
затратой энергии АТФ.
3Na+
2K+
АТФ
При действии на возбудимую ткань раздражителя подпороговой величины возникает местное возбуждение (локальный ответ), которое в отличие от
импульсного возбуждения:1.
Не подчиняется закону «все или ничего»
2.
Амплитуда ЛО зависит от силы стимула
3.
Распространяется по мембране затуханием
(декрементом)
4.
Может суммироваться (в результате амплитуда
деполяризации увеличивается)
5.
Трансформируется в потенциал действия при
достижении уровня критической деполяризации+30
0
Екр
Е0
1
4
5
3
2
Фазы возбудимости при генерации ПД
1.2.
3.
4.
5.
Повышенная возбудимость (соответствует
местному возбуждению)
Абсолютная рефрактерность – отсутствие
возбудимости (соответствует фазе
деполяризации и инверсии)
Относительная рефрактерность (фазе
реполяризации)
Супернормальная возбудимость (следовой
деполяризации)
Субнормальная возбудимость (следовой
гиперполяризации).
ЗАКОНЫ РАЗДРАЖЕНИЯ
Все возбудимые ткани подчиняютсяопределенным законам. К ним относятся:
закон силы
закон «все или ничего»
закон времени (длительности действия)
закон «крутизны» (времени нарастания
силы)
полярный закон
Законы раздражения
Закон силы – чтобы возник ПД, силавеличины. При этом ответная реакция
прямо пропорциональна силе раздражения.
Закон «все или ничего» - подпороговые
раздражители не вызывают ответной
реакции («ничего»), на пороговые и
сверпороговые – возникает максимальная
ответная реакция («всё»).
Законы раздражения
Закон времени – чтобы возник ПД, времядейстия стимула должно быть не меньше
пороговой величины. Чем больше сила
раздражения, тем меньше времени требуется,
чтобы произошло возбуждение ткани и наоборот,
чем меньше сила раздражения, тем больше
времени необходимо для возбуждения ткани.
Закон крутизны – чтобы возник ПД, крутизна
стимула должна быть не меньше пороговой
величины. При медленном нарастании силы
раздражения возбуждение может не возникнуть
(аккомодация).Аккомодация.
АккомодацияАккомодация проявляется в увеличении пороговой
силы стимула при уменьшении крутизны нарастании
стимула – чем меньше крутизна,
тем больше пороговая сила.
В основе аккомодации ткани лежит процесс
инактивации натриевых каналов.
Если крутизна нарастания стимула будет
меньше пороговой величины, то ПД не возникает
и будет наблюдаться только локальный ответ
(местное возбуждение).
Полярный закон
Сформулировал ученый Пфлюгер в 1859г.Существуют 3 положения полярного закона:
1. При действии постоянного тока возбуждение
возникает только в момент замыкания или только
в момент размыкания цепи.
2. При этом в момент замыкания возбуждение
возникает только под катодом, а в момент
размыкания - под анодом.
3. Возбуждение, которое возникает под катодом
больше, чем под анодом.Замыкание цепи
-+
+ катод
+
+
-
-
+
анод
-Размыкание цепи
- +
+ катод
-
+
анод
Полярный закон
Под катодом (отрицательным электродом)происходят
процессы
пассивной
деполяризации. При этом возбудимость ткани под
катодом повышается.
В то же время анод
(положительный электрод) вызывает пассивную
гиперполяризацию ткани. Возбудимость ткани под
анодом резко снижается. Это нашло применение в
медицинской
практике:
если
требуется
заблокировать проведение возбуждения по болевым
проводникам, то можно использовать постоянный
ток (анодный блок проведения возбуждения).
Физиологический электротон
Это изменения возбудимости мембраныпри воздействии на нее постоянного тока
подпороговой силы.
При этом под катодом развивается
катэлектротон
увеличение
возбудимости.
под анодом – анэлектротон - снижение
возбудимости.Электротон. А – катэлектротон.
1 – начальное повышение возбудимости: V1 < V.
2 – катодическая депрессия: V2 > V.
Б – анэлектротон, понижение возбудимости: V1 >
V.
Катодическая депрессия по Вериго
Если потоянный ток действует на мембранудлительное время, то повышенная возбудимость под катодом изменяется на снижение
возбудимости.
В основе этого явления лежит явление
аккомодации ткани, т.к. постоянный ток
можно представить как ток с бесконечно
малой крутизной нарастания.
Все живые клетки обладают раздражимостью, т.е. способностью реагировать на различные стимулы и переходить из состояния физиологического покоя в состояние активности. Этот процесс сопровождается изменением обмена веществ, а дифференцированные ткани (нервная, мышечная, железистая), осуществляющие специфические функции (проведение нервного импульса, сокращение или выделение секрета), - еще и изменением электрического потенциала.
Клетки возбудимых тканей могут находиться в трех различных состояниях (рис. 2.1). При этом клетки из состояния физиологического покоя могут переходить в активные состояния возбуждения или торможения, и наоборот. Клетки, находящиеся в состоянии возбуждения, могут переходить в состояние торможения, а из состояния торможения - в состояние возбуждения. Скорость перехода различных клеток или тканей из одного состояния в другое значительно различается. Так, двигательные нейроны спинного мозга могут от 200 до 300 раз в секунду переходить из состояния покоя в состояние возбуждения, тогда как вставочные нейроны - до 1000 раз.
Рис. 2.1.
Физиологический покой - состояние, характеризующееся:
- относительно постоянным уровнем обмена процессов;
- отсутствием функциональных проявлений ткани.
Активное состояние возникает под действием раздражителя и характеризуется:
- выраженным изменением уровня обменных процессов;
- проявлениями функциональных отправлений ткани. Возбуждение - активный физиологический процесс, возникающий под действием раздражителя, способствующий переходу ткани из состояния физиологического покоя к специфической деятельности (генерация нервного импульса, сокращение, секреция). Неспецифические признаки возбуждения:
- изменение заряда мембраны;
- повышение обменных процессов;
- увеличение затраты энергии.
Торможение - активный физиологический процесс, возникающий под действием определенного раздражителя и характеризующийся угнетением или прекращением функциональной активности ткани. Неспецифические признаки торможения:
- изменение проницаемости клеточной мембраны;
- изменение движения ионов через нее;
- изменение заряда мембраны;
- снижение уровня обменных процессов;
- снижение затраты энергии.
Раздражители, их классификация. Переход клеток из состояния физиологического покоя в состояние активности осуществляется под влиянием определенных факторов внешней или внутренней среды, так называемых раздражителей.
Раздражитель - это любое воздействие (вид энергии), способное вызвать биологическую реакцию живой ткани, изменение ее структуры и функции.
Различают внешние и внутренние раздражители. Внешние раздражители - разнообразные изменения окружающего мира - световые и звуковые волны, химические и механические воздействия на клетки. Внутренние раздражители - изменения состава и физико-химических свойств жидких сред организма, а также степени наполнения полых органов. Раздражители различают также по виду энергии. Выделяют химические, физические и биологические раздражители, например изменения pH, концентрации ионов, механические, температурные, электрические и др. Кроме того, раздражители различают по силе, длительности и характеру воздействия, физиологическому значению (адекватные и неадекватные) и другим признакам. Клетки более чувствительны к адекватным раздражителям, к восприятию которых они приспособились в процессе эволюции (например, свет - адекватный раздражитель для фоторецепторов, недостаток кислорода в артериальной крови - раздражитель для аортальных и каротидных хеморецепторов).
Наиболее часто при изучении свойств различных клеток и тканей в качестве раздражителя используют электрический ток, который называют универсальным раздражителем. Это обусловлено следующими причинами:
- электрический ток (до определенной силы) не оказывает на живую ткань необратимого влияния;
- электрический ток как раздражитель может быть точно градуирован по силе, длительности и градиенту своего воздействия на живую ткань (рис. 2.2);
- электрический ток близок к естественным механизмам возникновения и распространения возбуждения в живых тканях.

Рис. 2.2.
А - по силе; Б - длительности; В - градиенту
Величина ответной реакции клетки или ткани зависит от силы действующего раздражителя: чем сильнее раздражитель, тем сильнее (до известных пределов) и ответная реакция ткани.
На рис. 2.3 приведена зависимость между силой раздражителя и ответной реакцией ткани.

Рис. 2.3.
А - допороговые раздражители; Б - пороговый раздражитель;
В - субмаксимальный раздражитель; Г - максимальный раздражитель;
Д - супермаксимальный раздражитель
Как видно, слабые раздражители не вызывают видимой ответной реакции ткани. Такие раздражители принято называть подпороговыми. Отсутствие внешних признаков реагирования ткани (например, сокрашение мышц) не означает, что в клетках не происходит изменений обмена веществ и электрических процессов. Однако величина этих изменений (при действии подпорогового раздражителя) недостаточна для осуществления специфической функции клеток ткани.
Для проявления специфической функции ткани необходимо, чтобы воздействующий раздражитель имел определенную силу, равную или превышающую известную критическую величину. Такой раздражитель называют пороговым (рис. 2.4)". Раздражители, имеющие силу больше порогового, называют надпороговыми или субмаксимальными. При их воздействии величина ответа ткани возрастает до некоторого предела. Минимальный по силе раздражитель, вызывающий наибольший ответ ткани, называется максимальным раздражителем. Раздражители, сила которых превосходит силу максимальных раздражителей, называют супермаксимальными раздражителями. Все раздражители, дающие максимальный ответ, называют оптимальными. Раздражители, большие по величине, чем оптимальные, но вызывающие меньший ответ, чем при оптимальном раздражении, называют пессимальными.

Рис. 2.4.
Ответная реакция мышцы при действии раздражителей: 1 - подпорогового; 2 - порогового; 3 - субмаксимального; 4 - максимального; 5 - оптимального; 6 - пес- симального; 7 - супермаксимального; 8 - надпорогового
Основные свойства возбудимых тканей. Любая живая ткань обладает следующими свойствами: возбудимостью, проводимостью и лабильностью.
Возбудимость - способность ткани отвечать на действие раздражителей переходом в активное состояние. Возбудимость характерна для нервной, мышечной и железистой тканей. Возбудимость обратно
1 См.: Леонтьева Н.Н., Маринова К.В. Анатомия и физиология детского организма (основы учения о клетке и развитии организма, нервная система, опорно-двигательный аппарат). М.: Просвещение, 1986.
пропорциональна силе действующего раздражителя: В = 1/S. Чем больше сила действующего раздражителя, тем меньше возбудимость, и наоборот. Возбудимость зависит от состояния обменных процессов и заряда клеточной мембраны. Невозбудимостъ = рефрактерности Наибольшей возбудимостью обладает нервная ткань, затем поперечно-полосатая скелетная и сердечная мышечная ткань, железистая ткань.
Проводимость - способность ткани проводить возбуждение в двух или одном направлении. Показателем проводимости является скорость проведения возбуждения (от 0,5 до 120 м/с в зависимости от ткани и строения волокна). Быстрее всего возбуждение передается по миелинизированному нервному волокну, затем по немиелинези- рованному волокну, и самой низкой проводимостью обладает синапс.
Функциональная лабильность - способность ткани воспроизводить без искажения частоту ритмически наносимых импульсов. Показателем функциональной лабильности является количество импульсов, которое данная структура может передавать без искажения за единицу времени. Например, нерв - 500-1000 имп/с, мышца - 200-250 имп/с, синапс - 100-120 имп/с.
Роль силы раздражителя и времени его действия. Хронаксия - это временная характеристика возбудимости. Зависимость между пороговой интенсивностью раздражения и длительностью называют кривой силы длительности или кривой Гоорвега - Вейсса (рис. 2.5). Она имеет форму равносторонней гиперболы. На оси абсцисс откладывают время, на оси ординат - пороговую интенсивность раздражения.

Рис. 2.5.
По оси абсцисс отложено время (t); по оси ординат - пороговая интенсивность раздражения (/); 0А - реобаза; 0В-двойная реобаза; ОД - хропаксия; 0Ж-полезное время
Из рис. 2.5 можно видеть, что при слишком малой величине интенсивности раздражения (менее ОА) ответная реакция не возникает при любой его длительности. Отсутствует реакция и при слишком малом времени действия раздражителя (менее ОГ). При интенсивности раздражения, соответствующей отрезку ОА, возникает возбуждение при условии большей длительности действия раздражающего импульса. В пределах времени, определяемого отрезком ОЖ, имеет место зависимость между пороговой интенсивностью и длительностью раздражения: меньшей длительности раздражающего импульса соответствует большая пороговая интенсивность (отрезку ОД соответствует OB, а ОЕ - отрезку ОБ). За пределами этого времени (ОЖ) изменение длительности действия раздражителя уже не влияет на величину порога раздражения. Наименьшее время, в течение которого проявляется зависимость между пороговой интенсивностью раздражения и его длительностью, получило название полезного времени (отрезок ОЖ). Полезное время является временным показателем возбуждения. По его величине можно судить о функциональном состоянии различных возбудимых образований. Однако для определения полезного времени необходимо найти несколько точек кривой, для чего требуется наносить множество раздражений. Поэтому большое распространение получило определение другого временного показателя, который ввел в практику физиологических исследований Л. Лапик (1907). Он предложил для характеристики скорости возникновения процесса возбуждения параметры: реобазу и хронаксию.
Реобаза - это пороговая интенсивность раздражения при большой длительности его действия (отрезок ОА); хронаксия - время, в течение которого должен действовать ток, равный двойной реобазе (ОВ), для получения порогового ответа (отрезок ОД). В течение этого времени происходит уменьшение мембранного потенциала до величины, соответствующей критическому уровню деполяризации. Для разных возбудимых образований величина хронаксии неодинакова. Так, хронаксия локтевого нерва человека составляет 0,36 мс, срединного - 0,26 мс, общего сгибателя пальцев - 0,22 мс, а общего разгибателя - 0,58 мс.
- См.: Леонтьева Н.Н., Маринова К.В. Указ. соч.